Introduction
In 2020, breast cancer surpassed lung cancer as the most common cancer worldwide, accounting for 12.5% of cancer diagnoses [1]. The incidence of breast cancer continues to increase, with a projected increase of > 40% in new cases and > 50% in deaths by 2040 [1]. Approximately 88% of patients with breast cancer undergo at least one anesthetic and surgical treatment within one year of diagnosis [2]. However, surgery and anesthesia are associated with an increased release of inflammatory mediators and angiogenic factors, and cause postoperative immunosuppression, resulting in tumor progression [3].
The natural killer group 2, member D (NKG2D) ligands, UL16-binding proteins (ULBP) 1–3, and major histocompatibility complex class I chain-related molecules (MIC) A/B, which are expressed on the surface of cancer cells, bind to active receptors on natural killer cells (NK cells), transmitting signals and allowing NK cells to recognize and eliminate cancer cells [4]. A reduction in the expression of NKG2D ligands can impair the cytotoxicity of NK cells against cancer cells, leading to immune evasion and disruption of the cancer immunosurveillance system [5].
Matrix metalloproteinases (MMPs) are Zn2+-dependent endopeptidases that play a critical role in tumor progression by promoting extracellular matrix and basement membrane degradation, leading to cell detachment and migration [5,6]. MMPs also promote neovascularization and contribute to tumor angiogenesis [5,6]. Moreover, recent cancer immunological studies have demonstrated that MMPs cleave and remove NKG2D ligands [5–7]. MMP-induced shedding of NKG2D ligands confers several advantages to cancer cells for immune evasion [8,9]. First, it reduces the density of NKG2D ligands on the surface of cancer cells, thereby impairing their susceptibility to NK cells [8,9]. Moreover, the cleaved ligands (soluble NKG2D ligands) retain their ability to bind to the NKG2D receptors on NK cells [8,9]. This cleaved ligand-receptor engagement not only hinders the activation signaling of the receptor, but also triggers the internalization and downmodulation of the NKG2D receptor on NK cells [8,9].
The modulation of surgery-related factors in clinical practice remains challenging and requires an enhanced understanding of the effects of anesthesia-related factors on cancer recurrence and survival rates. Therefore, elucidating the effects of anesthetics on the breast cancer microenvironment is essential for optimal anesthesia management and an improvement in postoperative outcomes.
In this study, we investigated the effects of sevoflurane, a common general anesthetic, on MMP expression and NKG2D-mediated NK cell cytotoxicity in breast cancer cells. We evaluated the effects of sevoflurane on MMP expression, NKG2D ligand expression and ablation, and NK cell-mediated cytotoxicity in breast cancer cells.
Materials and Methods
Cell lines and reagents
This study was conducted using the following three breast cancer cell lines: estrogen receptor (ER)- and progesterone receptor (PR)-positive human breast cancer cell line MCF-7 (Korean Cell Line Bank, Korea), human epidermal growth factor receptor 2 (HER2)-positive human breast cancer cell line MDA-MB-453 (Korean Cell Line Bank), and triple-negative human breast cancer cell line HCC-70 (Korean Cell Line Bank) [10]. All cell lines were maintained in RPMI-1640 medium supplemented with 10% fetal bovine serum and 1% penicillin (Welgene, Korea). Because sevoflurane is a strong organic solvent capable of interacting with plastic materials to produce impurities [11], breast cancer cells were cultured in poly-L-lysine-coated (Sigma-Aldrich, USA) glass culture dishes.
The human NK cell line, NK92-MI (ATCC, USA), was maintained in an α-minimum essential medium containing fetal bovine serum (12.5%), horse serum (12.5%), 2 mercaptoethanol (0.1 mM), and L-glutamine (2 mM). All cell lines were cultured according to their specifications and incubated at 37°C in humidified air containing 5% CO2.
Sevoflurane treatment
As previously described [12–14], 100 μl sevoflurane (Sevoprane; Ilsung, Korea) was diluted in 10 ml RPMI-1640 medium and stirred for a half-hour in an airtight, amber-colored glass bottle. The concentration of sevoflurane was determined using the preliminary data obtained from our gas chromatography-mass spectrometry analysis (GCMS-QP2010 Plus; Shimadzu, Japan) as (mean [SD]): 3.92 (1.26) mM. The sevoflurane stock was serially diluted to 1200 and 600 µM (S12 and S6, respectively) immediately before the experiments. Kharasch et al. [15] simultaneously measured the end-tidal sevoflurane concentration and plasma concentration of sevoflurane during general anesthesia and revealed that the average peak plasma concentration of sevoflurane reached 772 μM at an end-tidal sevoflurane concentration of 2.7% (equivalent to 1.3 minimum alveolar concentration). Consequently, we assumed clinically relevant concentrations of sevoflurane at 600 μM during general anesthesia. Furthermore, to investigate the dose-response relationship, we administered an additional dose of sevoflurane at a concentration of 1200 μM, which was higher than the recommended dose. The corresponding concentration of distilled water in the RPMI 1640 media were used as controls (0 μM).
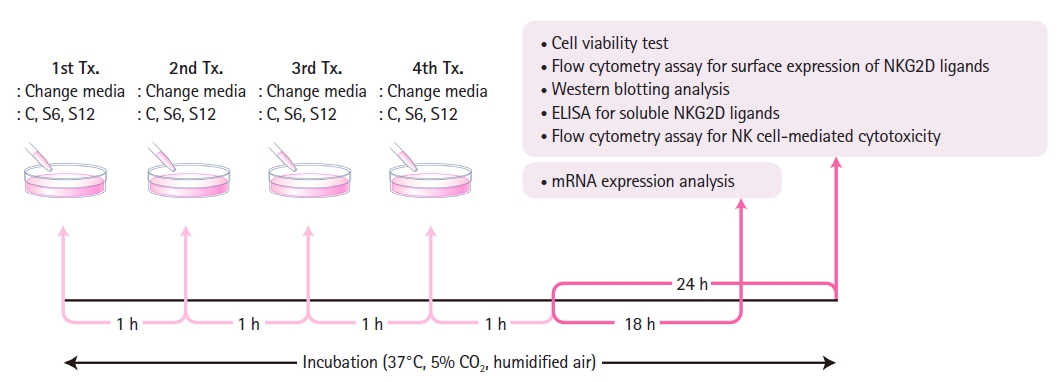
The MCF-7, MDA-MB-453, and HCC-70 cells were then exposed to sevoflurane for 4 h. To account for evaporation-induced concentration reduction, both the sevoflurane and control group solutions were replaced every hour (Fig. 1) [13,14]. Previous studies [12,13] have shown that despite the volatility of sevoflurane, the concentration of the sevoflurane solution dissolved in cell culture media remains stable, with a < 10% loss over a one-hour period.
mRNA expression analysis was performed 18 h after treatment completion, while the other experiments (cell viability test, flow cytometry assay for surface expression of NKG2D ligands, western blotting analysis, enzyme-linked immunosorbent assay [ELISA] for soluble NKG2D ligands, and flow cytometry assay for NK cell-mediated cytotoxicity) were performed 24 h after treatment completion (Fig. 1).
Cell viability test
The MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide) assay is based on the principle that mitochondrial activity in living cells converts MTT into formazan crystals. These crystals are dissolved following the addition of dimethyl sulfoxide (DMSO) and detected spectrophotometrically at 540 nm, with the absorbance directly proportional to cell viability [16]. As previously described [14], the MCF-7, MDA-MB-453, and HCC-70 cells were plated in 96-well plates (1 × 104 cells/well) and incubated with the control or S12 solution for 4 h. Twenty-four hours later, the cells were incubated with the MTT solution (Sigma-Aldrich) for 4 h. The supernatant was discarded and the formazan crystals were dissolved using DMSO. The absorbance at 540 nm was determined using a microplate spectrophotometer (μQuant; Bio Tek, USA).
mRNA expression analysis of NKG2D ligands
The mRNA expression analysis method used has been described previously [14]. Briefly, after a 4-h treatment and an additional 18-h incubation period, cancer cells were harvested (Fig. 1). The total RNA was extracted from the cells using a RNeasy®Mini kit (Qiagen GmbH, Germany). Reverse transcription polymerase chain reaction (RTPCR) and multiplex PCR were performed. For denaturation, 3 μg extracted total RNA and 100 pmol random primers (Takara Shuzo, Japan) were incubated at 65°C for 5 min and chilled at 4°C for 4 min. Next, 6 μl of the 5x reaction buffer, 4 μl deoxynucleotide triphosphate (10 mM; Promega Co., USA), and 1.2 μl M-MLV RT (Promega Co.) were added and incubated at 37°C for 60 minutes. Multiplex PCR was performed using a QIAGEN® Multiplex PCR kit (Qiagen GmbH). The primer sets used to evaluate NKG2D gene expression were as follows: 1) MICA: ribosomal protein L19 (RPL19), MICA, and β-actin genes and 2) MICB and ULBP 1–3: RPL19, MICB, ULBP1–3, and β-actin genes. The primer sequences are listed in Table 1. All the experiments were performed according to the manufacturer’s instructions. PCR products were quantified using a microchip electrophoresis system MCE®-202MultiNA (Shimadzu, Japan). For normalization, the mRNA band intensity of each NKG2D ligand was divided by that of the β-actin. To assess relative gene expression ratios, the normalized mRNA band intensity of the treated samples was divided by that of the controls.
Flow cytometry assay for surface expression of NKG2D ligands
The flow cytometry assay for the surface expression of NKG2D ligands was performed using the method described in our previous study [14]. Briefly, after a 4-h treatment and an additional 24-h incubation period (Fig. 1), the cells were harvested and incubated with 10 μg/ml mouse anti-MICA/B and ULBP1–3 or the corresponding isotype controls (anti-IgG2a or anti-IgG2b; R&D Systems, USA). Samples were then incubated with goat anti-mouse phycoerythrin (PE)-conjugated antibodies (BD Biosciences, USA). The mean fluorescence intensity (MFI) was measured using a FACSCanto II flow cytometer (BD Biosciences) and quantified using FlowJo (version 10.6.1; TreeStar, USA). The MFIs of the treated samples were divided by those of the controls to assess the relative expression ratios.
Western blot analysis for determining protein expression of MMPs
After a 4-h treatment and an additional 24-h incubation period (Fig. 1), western blot analysis was performed to evaluate MMP-1 and -2 expression. The cells were washed three times with cold phosphate-buffered saline and lysed in a PRO-PREP protein extraction solution (Intron, Korea). Equal amounts of cell extracts were resolved by 4%–20% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and analyzed by western blotting. The separated proteins were transferred onto polyvinylidene difluoride membranes (Millipore, USA). The membranes were then blocked with 3% BSA in Tris-buffered saline containing 0.1% Tween 20 at room temperature. The proteins of interest were detected using primary antibodies (MMP-1 and -2; Cell Signalling, USA) and horseradish peroxidase-conjugated secondary antibodies (Enzo Life Sciences, USA) according to the manufacturer’s instructions, and a chemiluminescence imaging system (AE-9150 Ez-capture II; Atto, Japan) was used to analyze the results. Each blot was probed with anti-β-actin antibodies (Sigma-Aldrich). Band intensity was quantified using ImageJ software (version 1.53; National Institutes of Health, USA). Protein expression in the treated cells was divided by that in the control cells to calculate relative protein expression ratios.
Enzyme-linked immunosorbent assay (ELISA) for soluble NKG2D ligands
Breast cancer cells (4 × 106 cells) were plated on 60-mm glass culture dishes. After a 4-h treatment and an additional 24-h incubation period (Fig. 1), cell culture supernatants were centrifuged at 5000 rpm for 5 min at 4°C, and aliquots were stored at −80°C until further use. The levels of soluble NKG2D ligands (MICA for MCF-7 and MDA-MB-453; MICB for HCC-70) in the cell culture supernatant were measured using ELISA kits (MICA Human ELISA kit, Invitrogen, USA; Human MICB ELISA kit, MyBioSource, USA) following the manufacturer’s protocol. The absorbance at 450 nm was measured using a microplate spectrophotometer (Synergy H1; BioTek, USA). The samples were loaded in duplicate, and the mean soluble NKG2D ligand values were used for analysis.
Flow cytometry assay for NK cell-mediated cytotoxicity
After a 4-h treatment and an additional 24-h incubation period (Fig. 1), target cancer cells (MCF-7, MDA-MB-453, and HCC-70) were stained with carboxyfluorescein diacetate succinimidyl ester (CFSE; CellTraceTM; Invitrogen) and co-cultured with NK92-MI cells, effector cells, for 4 h. With cellular cytotoxicity assays, optimizing the effector-to-target cell (E:T) ratio is crucial for maximizing and distinguishing differences in cytotoxicity among treatment groups. Typically, higher E:T ratios result in enhanced NK cell cytotoxicity because an increased number of effector cells relative to target cells leads to greater cytotoxicity [17]. The recommended E:T ratio for flow cytometry cytotoxicity assays is ≤ 10:1 [18]. As previously reported [14], we determined the E:T ratio to be between 1:1 (e.g., E:T = 1 × 105 : 1 × 105) and 10:1 (e.g., E:T = 1 × 106 : 1 × 105), with 10:1 as the optimal ratio. Co-cultured cells were labelled with 1 μg/ml propidium iodide (PI; Invitrogen). A FACSCantoTM II flow cytometer and BD FACSDivaTM Software (BD Biosciences) were used. The percentage of NK cell-mediated lysis (%) was calculated using the following equation:
where Q2 represents CFSE-positive and PI-positive cells and Q3 represents CFSE-positive and PI-negative cells.
Statistical analysis
MedCalc® (version 20; MedCalc Ltd., Belgium) and IBM SPSS Statistics (version 25; IBM Corp., USA) were used for statistical analyses. Variables are presented as medians with the first and third quartiles (Q1, Q3). For comparisons between groups, Mann-Whitney U tests or Kruskal-Wallis tests were performed. If the Kruskal-Wallis test was significant, post-hoc comparisons using the Conover method were conducted.
Our trial consisted of a zero-dose control group (C) and two sevoflurane treatment groups (S6 and S12) that received increasing doses of sevoflurane in the following order: C, S6, S12. As a secondary outcome, the dose-response relationship was evaluated using the Jonckheere-Terpstra trend test. Statistical significance was set at P < 0.05.
Results
Effect of sevoflurane on cell viability assessed by MTT assay
Sevoflurane was not found to affect the viability of MCF-7, MDA-MB-453, or HCC-70 cells. The relative cell viabilities (%) related to the controls measured by MTT assay were as follows (median [Q1, Q3]): MCF-7 at S12: 100.2 (92.4, 118.9), P = 1.000; MDA-MB-453 at S12: 101.4 (96.2, 109.0), P = 1.000; and HCC-70 at S12: 89.4 (87.3, 107.0), P = 0.305 (n = 6 per group).
Effect of sevoflurane on the mRNA expression of NKG2D ligands
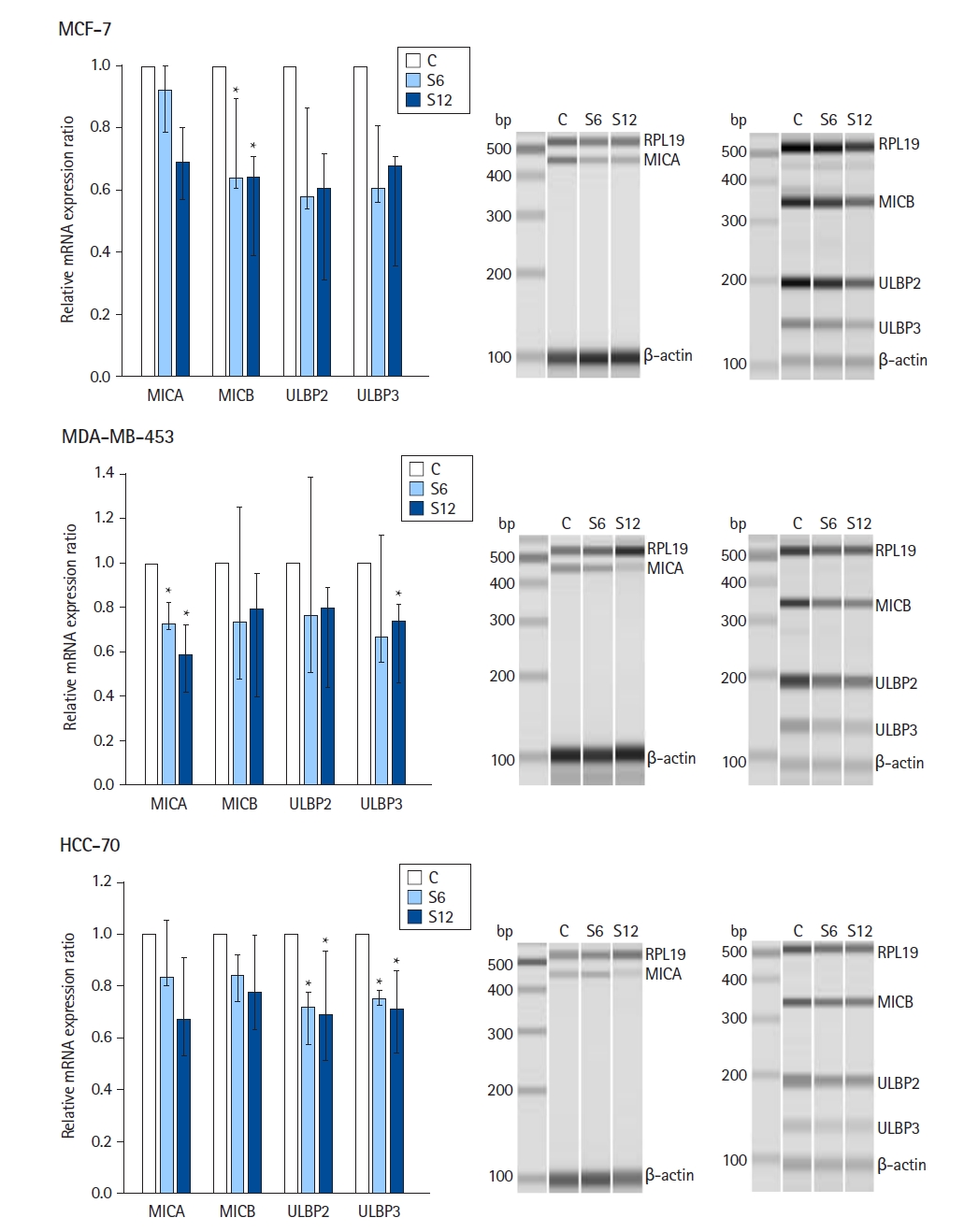
The results of sevoflurane exposure on the mRNA expression of NKG2D ligands are summarized in Fig. 2 (n = 6 per group). In MCF-7, MDA-MB-453, and HCC-70 cell lines, mRNA expression of MICA, MICB, ULBP2, and ULBP3 was observed, but the expression of ULBP1 was not observed.
In MCF-7 cells, the relative mRNA expression ratios of MICB at S6 and S12 were lower than those in the controls (Kruskal-Wallis test: P = 0.013). In MDA-MB-453 cells, the relative mRNA expression ratios of MICA at S6 and S12 were lower than those in the controls (Kruskal-Wallis test: P = 0.013) and the relative surface expression ratio of ULBP3 was downregulated at S12 compared with the controls (Kruskal-Wallis test: P = 0.044). In HCC-70 cells, the relative mRNA expression ratios of ULBP2 and ULBP3 at S6 and S12 were lower than those in the controls (Kruskal-Wallis test: P = 0.002 and P = 0.003, respectively).
The secondary outcome results showed a dose-response relationship in MCF-7 cells, demonstrating significantly lower relative mRNA expression levels of MICA, MICB, ULBP2, and ULBP3 in response to increasing concentrations of sevoflurane (Jonckheere-Terpstra trend test: P = 0.015, P = 0.008, P = 0.036, and P = 0.043, respectively). Similarly, sevoflurane downregulated the mRNA expression of MICA and ULBP3 in MDA-MB-453 cells in a dose-dependent manner (Jonckheere-Terpstra trend test: P = 0.002 and P = 0.029, respectively). Likewise, sevoflurane downregulated the mRNA expression of MICA, MICB, ULBP2, and ULBP3 in HCC-70 cells in a dose-dependent manner (Jonckheere-Terpstra trend test: P = 0.043, P = 0.043, P = 0.006, and P = 0.012, respectively).
Effect of sevoflurane on surface expressions of NKG2D ligands assessed by flow cytometry
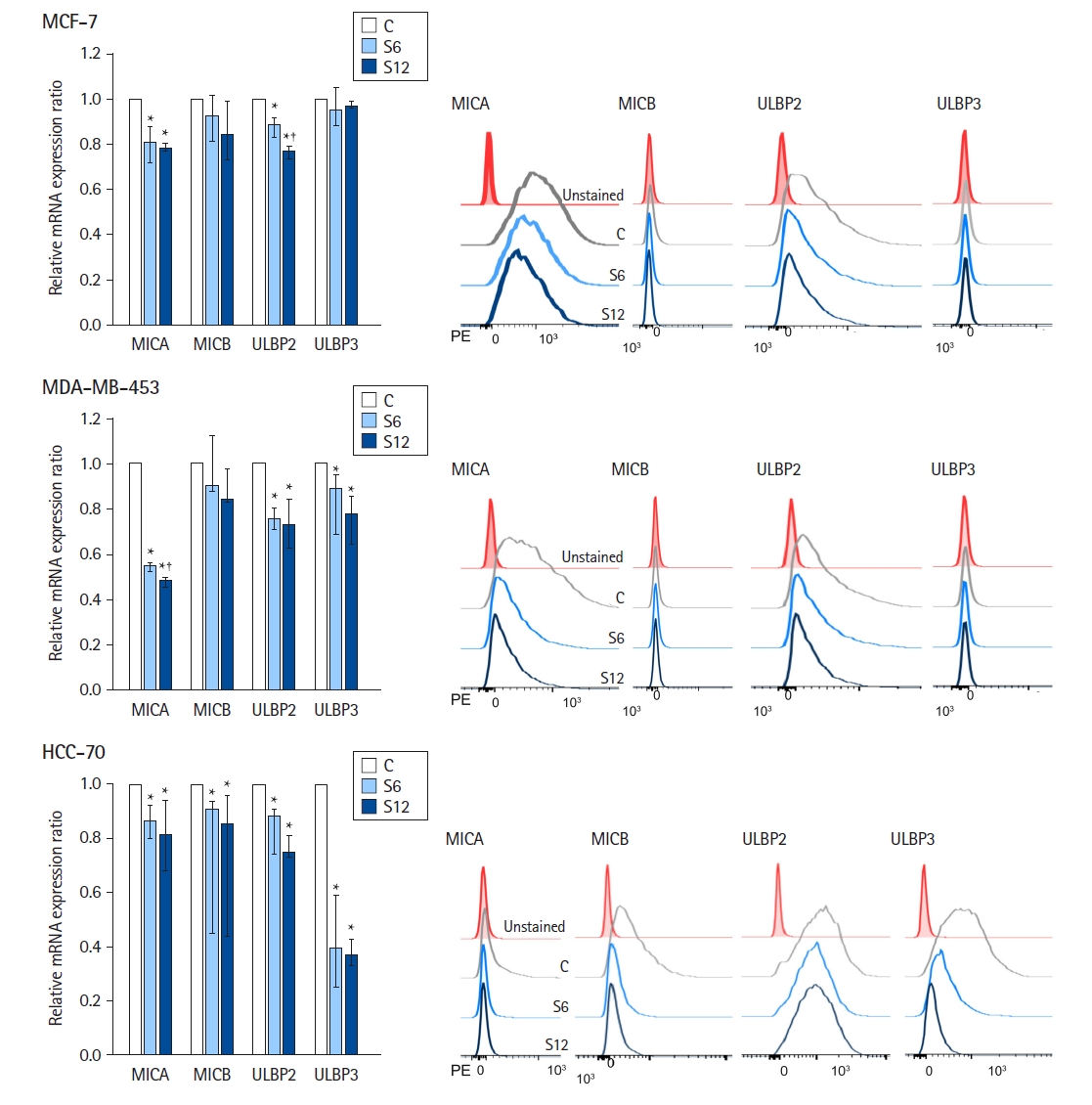
Fig. 3 summarizes the results of the flow cytometry analysis of NKG2D ligand surface expression (n = 6 per group). Consistent with mRNA expression, ULBP1 was rarely expressed on the surface of the MCF-7, MDA-MB-453, and HCC-70 cell lines.
In MCF-7 cells, MICA and ULBP2 were predominantly expressed; the relative surface expression ratios of MICA and ULBP2 at S6 and S12 were downregulated compared to those in the controls (Kruskal-Wallis test: P = 0.002 and P ≤ 0.001, respectively).
MDA-MB-453 cells predominantly exhibited surface expression of MICA and ULBP2. The relative surface expression ratios of MICA, ULBP2, and ULBP3 at S6 and S12 were lower than those in the controls (Kruskal-Wallis test: P < 0.001, P = 0.003, and P = 0.004, respectively).
In HCC-70 cells, MICB, ULBP2, and ULBP3 were predominantly expressed. The relative surface expression ratios of MICA, MICB, ULBP2, and ULBP3 at S6 and S12 were downregulated compared with those in the controls (Kruskal-Wallis test: P = 0.002, P = 0.017, P = 0.002, and P = 0.013, respectively).
In the dose-response analysis, we observed significantly lower relative surface expression ratios of MICA, MICB, and ULBP2 in MCF-7 cells with increasing concentrations of sevoflurane (Jonckheere-Terpstra trend test: P < 0.001, P = 0.024, and P < 0.001, respectively). Similarly, sevoflurane decreased the relative surface expression ratios of MICA, ULBP2, and ULBP3 in MDA-MB-453 cells in a dose-dependent manner (Jonckheere-Terpstra trend test: P < 0.001, P = 0.002, and P = 0.001, respectively). Likewise, sevoflurane downregulated the relative surface expression ratios of MICA, MICB, ULBP2, and ULBP3 in HCC-70 cells in a dose-dependent manner (Jonckheere-Terpstra trend test: P = 0.001, P = 0.010, P < 0.001, and P = 0.029, respectively).
Effect of sevoflurane on protein expression of MMP assessed by western blot analysis
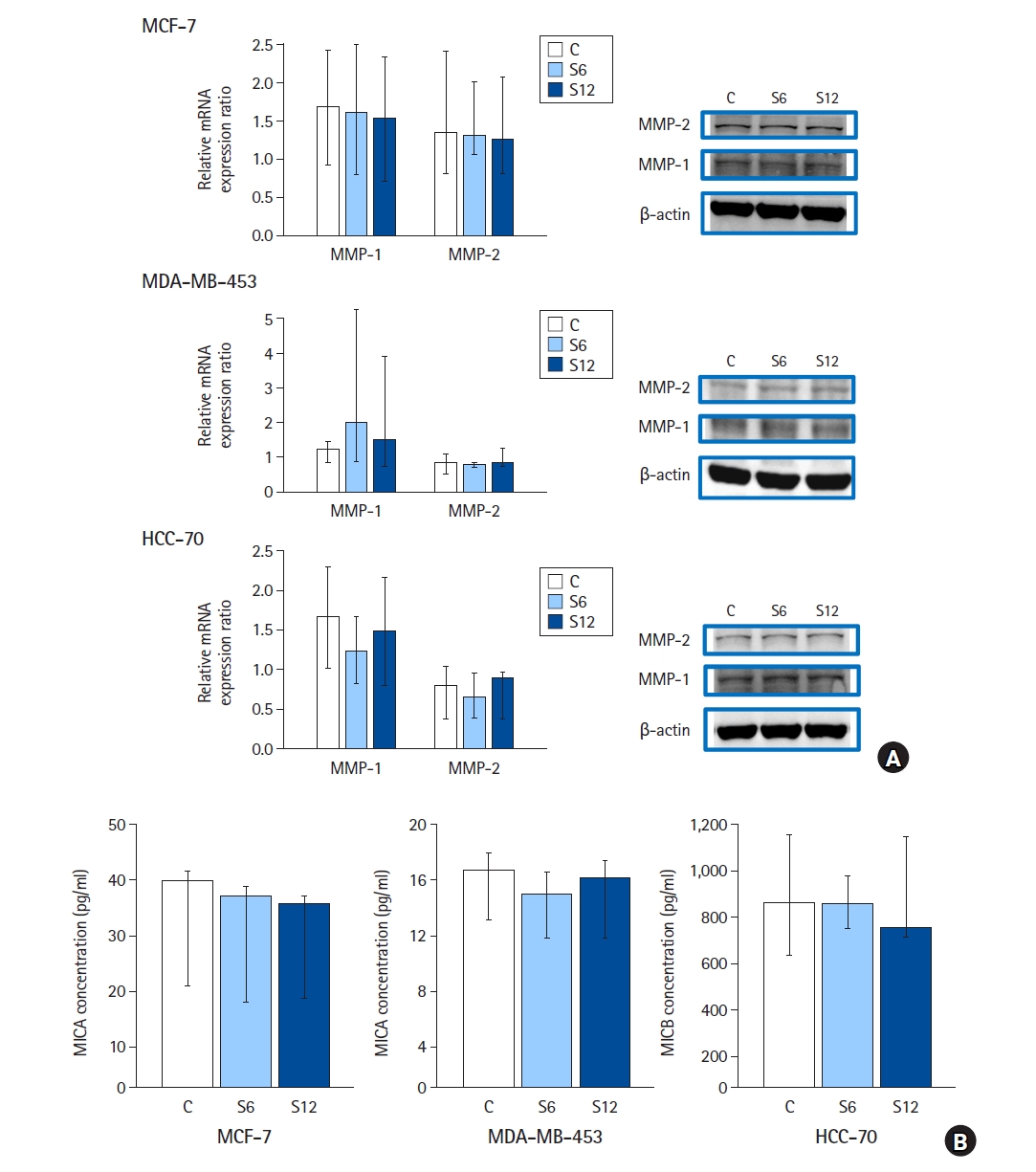
Western blot analysis revealed no changes in the protein expression of MMP-1 and -2 between the control and sevoflurane treatment groups in MCF-7, MDA-MB-453, and HCC-70 cells (Fig. 4A; n = 6 per group).
In MCF-7 cells, the median (Q1, Q3) of the relative protein expression ratio of MMP-1 was as follows: controls, 1.7 (0.9, 2.4); S6, 1.6 (0.8, 2.5); and S12, 1.5 (0.7, 2.3). P values for the Kruskal-Wallis and Jonckheere-Terpstra trend tests were 0.519 and 0.258, respectively. The median (Q1, Q3) of the relative protein expression ratio of MMP-2 was as follows: controls, 1.3 (0.8, 2.4); S6, 1.3 (1.1, 2.0); and S12, 1.3 (0.8, 2.1). P values for the Kruskal-Wallis and Jonckheere-Terpstra trend tests were 0.854 and 0.686, respectively.
In MDA-MB-453 cells, the median (Q1, Q3) of the relative protein expression ratio of MMP-1 was as follows: controls, 1.3 (0.9, 1.5); S6, 2.0 (0.9, 5.3); and S12, 1.5 (0.7, 3.9). P-values for the Kruskal-Wallis and Jonckheere-Terpstra trend tests were 0.653 and 0.628, respectively. The median (Q1, Q3) of the relative protein expression ratio of MMP-2 was as follows: controls, 0.9 (0.5, 1.1); S6, 0.8 (0.7, 0.9); and S12, 0.8 (0.8, 1.3). P values for the Kruskal-Wallis and Jonckheere-Terpstra trend tests were 0.778 and 0.746, respectively.
In HCC-70 cells, the median (Q1, Q3) of the relative protein expression ratio of MMP-1 was as follows: controls, 1.7 (1.0, 2.3); S6, 1.2 (0.8, 1.7); and S12, 1.4 (0.8, 2.1). P-values for the Kruskal-Wallis and Jonckheere-Terpstra trend tests were 0.423 and 0.293, respectively. The median (Q1, Q3) of the relative protein expression ratio of MMP-2 was as follows: controls, 0.8 (0.4, 1.0); S6, 0.7 (0.4, 1.0); and S12, 0.9 (0.4, 1.0). P values for the Kruskal-Wallis and Jonckheere-Terpstra trend tests were 0.911 and 0.808, respectively.
Sevoflurane did not affect the soluble NKG2D ligand concentration assessed by ELISA
According to the ELISA results, no significant changes were detected in the levels of soluble NKG2D ligands between the control and sevoflurane-treated groups in MCF-7, MDA-MB-453, or HCC-70 cells (Fig. 4B, n = 6 per group).
According to the Jonckheere-Terpstra test results, no dose-response relationship was identified. P values for the MCF-7, MDA-MB-453, and HCC-70 cells were 0.467, 0.125, and 0.686, respectively.
Effect of sevoflurane on NK cell-mediated cytotoxicity assessed by flow cytometry
The flow cytometry results are shown in Fig. 5 (n = 4 per group). In all cell lines, NK cell-mediated cytotoxicity at S6 and S12 was lower than that in the controls. Compared with the controls, the P values were as follows: MCF-7 at S6: 0.005 and MCF-7 at S12: 0.005; MDA-MB-453 at S6: 0.005 and MDA-MB-453 at S12: 0.005; and HCC-70 at S6: 0.005 and HCC-70 at S12: 0.005 (Fig. 5; effect cells:target cells = 10:1, n = 4 per group).
Analysis of the secondary outcomes using the Jonckheere-Terpstra trend test revealed that sevoflurane reduced NK cell-mediated cytotoxicity in MCF-7, MDA-MB-453, and HCC-70 cells in a dose-dependent manner (effector cells:target cells = 10:1; P = 0.040, P = 0.040, and P = 0.040, respectively).
Discussion
In this study, we demonstrated that sevoflurane downregulated the mRNA and protein expression of NKG2D ligands in human breast cancer cell lines in a dose-dependent manner. However, sevoflurane was not found to affect the expression of MMP-1 and -2 or the concentration of proteolytically cleaved soluble NKG2D ligands. Furthermore, sevoflurane attenuated NK-cell-mediated cancer cell lysis in a dose-dependent manner.
The association between anesthetics and cancer recurrence was first reported in the 2000s. Since then, several preclinical and clinical studies have been conducted to identify the potential effects of anesthetics and anesthesia methods on breast cancer prognosis [19–21]. Although preclinical trials have suggested potential associations between anesthetic agents and breast cancer invasion and metastasis, the results of clinical research comparing these effects remain inconclusive [19]. A recent meta-analysis, in a subgroup analysis of breast cancer based on a prospective and three retrospective clinical studies, reported that recurrence-free survival and overall survival rates of breast cancers did not improve when total intravenous anesthesia was used compared with inhalation anesthesia (hazard ratio [HR], 95% CI of recurrence-free survival: 0.83 [0.59, 1.15]; overall survival: 1.12 [0.90, 1.39]) [22]. Sessler et al. [23] conducted a multicenter randomized controlled trial and demonstrated that recurrence-free survival did not differ between sevoflurane-based general and regional anesthesia with propofol (adjusted HR, 95% CI: 0.97 [0.74, 1.28]).
Our results demonstrate that sevoflurane suppresses NK cell-mediated cancer cell lysis in a dose-dependent manner. Consistent with our results, previous studies have demonstrated a potential association between sevoflurane exposure and the inhibition of NK cell activity and have suggested various underlying mechanisms in breast cancer patients [24,25]. In a pilot clinical trial conducted on ten breast cancer patients, sevoflurane-based general anesthesia reduced the expression of the NK cell-activating receptor (CD16) and their cytokines (interleukin [IL]-1β and IL-10) and decreased NK cell-mediated cytotoxicity [24]. Similarly, in another randomized controlled trial that included 50 participants, NK cell-mediated cytotoxicity decreased after breast cancer resection under sevoflurane-based anesthesia with fentanyl analgesia [25].
In our previous study on a non-small cell lung cancer cell line, sevoflurane administered at an anesthetic dose decreased NKG2D ligand expression and NK cell-mediated cytotoxicity. This effect was attributed to the suppression of NKG2D ligand transcription and an increase in MMP expression [14]. However, the present study on breast cancer cell lines suggests a different mechanism for the reduction of NK cell-mediated cytotoxicity by sevoflurane. We propose that sevoflurane directly inhibits the transcription of NKG2D ligands rather than the NKG2D ligand shedding induced by increased MMP expression.
Previous studies examining the effects of sevoflurane on MMP expression in breast cancer have yielded conflicting results [26,27]. Deegan et al. [26] reported that, following primary breast cancer surgery, sevoflurane with opioid anesthesia increased the serum levels of MMP-3 and MMP-9, but not MMP-1, compared to propofol with a paravertebral block. In contrast, Galos et al. [27] demonstrated no difference in the serum levels of MMP-3 and MMP-9 before or after sevoflurane-based anesthesia in patients with breast cancer. These discrepancies could be attributed to the heterogeneity in cancer subtypes, patient characteristics, and anesthetic exposure regimens. However, only a limited number of studies have evaluated the effects of anesthetic agents on NK cell ligand expression in breast cancer.
Our study had a few limitations. First, as this was an in vitro study, our results are not directly applicable to animals or humans. Second, the present study was not designed to elucidate the molecular mechanism by which sevoflurane affects the expression of NKG2D ligands and MMPs; therefore, further studies are warranted to understand the detailed mechanism. Third, sevoflurane was the only anesthetic agent used in this study. Consequently, whether our results represent a universal phenomenon associated with higher concentrations of anesthetics or are specific to sevoflurane remains unknown. For a comprehensive understanding of our findings, additional research using other anesthetic agents such as propofol is necessary.
In summary, sevoflurane was found to attenuate NK cell-mediated cancer cell lysis in a dose-dependent manner, which could be attributed to the sevoflurane-induced decrease in the transcription of NKG2D ligands rather than sevoflurane-induced changes in MMP expression and their proteolytic activity. Further research is essential to elucidate the effects of sevoflurane on immune escape and immunosurveillance in breast cancer.