Effects of remote ischemic postconditioning on hepatic injury in lipopolysaccharide-induced endotoxemic rats
Article information

Abstract
Background
Remote ischemic postconditioning (RIPoC) is induced by several cycles of brief, reversible, mechanical blood flow occlusion, and reperfusion of the distal organs thereby protecting target organs. We investigated if RIPoC ameliorated liver injury in a lipopolysaccharide (LPS)-induced endotoxemic rats.
Methods
Protocol 1) Rats were administered LPS and samples collected at 0, 2, 6, 12, and 18 h. 2) After RIPoC at 2, 6, and 12 h (L+2R+18H, L+6R+18H, and L+12R+18H), samples were analyzed at 18 h. 3) RIPoC was performed at 2 h, analysis samples at 6, 12, 18 h (L+2R+6H, L+2R+12H, L+2R+18H), and RIPoC at 6 h, analysis at 12 h (L+6R+12H). 4) Rats were assigned to a control group while in the RIPoC group, RIPoC was performed at 2, 6, 10, and 14 h, with samples analyzed at 18 h.
Results
Protocol 1) Liver enzyme, malondialdehyde (MDA), tumor necrosis factor-α (TNF-α), and nuclear factor-κB (NF-κB) levels increased while superoxide dismutase (SOD) levels decreased over time. 2) Liver enzyme and MDA levels were lower while SOD levels were higher in L+12R+18H and L+6R+18H groups when compared with L+2R+18H group. 3) Liver enzyme and MDA levels were lower while SOD levels were higher in L+2R+6H and L+6R+12H groups when compared with L+2R+12H and L+2R+18H groups. 4) Liver enzyme, MDA, TNF-α, and NF-κB levels were lower while SOD levels were higher in RIPoC group when compared with control group.
Conclusions
RIPoC attenuated liver injury in the LPS-induced sepsis model by modifying inflammatory and oxidative stress response for a limited period.
Introduction
Sepsis is a systemic inflammatory response to infection and generates harmful consequences, such as multiple organ damage and death. Despite significant improvements in its treatment, sepsis still accounts for 26.7% and 41.9% of all deaths in hospitals and intensive care units, respectively [1]. Primary sepsis treatment involves source control with antibiotics; however, in recent years, modulating host inflammatory responses (i.e., using corticosteroids, prostaglandins, and leukotriene inhibitors) has garnered considerable research interest. Although several novel treatment modalities have been introduced as prospective sepsis treatment options, none are definitive and thus further research is required [2].
The pathophysiology of sepsis-induced multi-organ damage is extremely complex. Uncontrolled systemic inflammatory responses and ischemia/reperfusion (I/R) processes related to low cardiac output, arterial hypotension, microcirculation failure, and mitochondrial dysfunction are believed to be contributing factors [3–5]. Specifically, the liver contributes to inflammatory responses by generating proinflammatory cytokines and acute phase proteins [6,7]. Therapeutic interventions reducing sepsis-induced inflammatory cytokines and hepatic dysfunction caused by cytokines may reduce multiple organ damage and mortality risks in sepsis [8].
Remote ischemic conditioning (RIC) is manifested by several cycles of brief, reversible, and non-lethal mechanical blood flow occlusion of distal tissue or organs, with reperfusion exerting protective effects on distant target tissue or organs. RIC can be performed either before ischemia (remote ischemic preconditioning [RIPC]) or after ischemia (remote ischemic postconditioning [RIPoC]) [9]. Several clinical trials have suggested that RIC is a promising approach that protects multiple organs (e.g., brain, heart, and kidneys) against the deleterious effects of I/R, even though inconsistencies exist in the literature [10–12]. Although the precise mechanisms underpinning these protective effects remain unclear, the effects are associated with downregulated systemic inflammatory and oxidative stress responses. Although few studies have investigated the effects of RIPC or RIPoC in sepsis, the application and therapeutic efficacy of RIPoC toward hepatic dysfunction in sepsis remain unknown.
Therefore, to address this, we examined if RIPoC ameliorated liver injury via anti-inflammatory and antioxidant marker levels in a lipopolysaccharide (LPS)-induced sepsis rat model.
Materials and Methods
Ethics statement
The Kyungpook National University Institutional Animal Care and Use Committee approved all experimental protocols (approval No: 2021-0159). Procedures were executed in accordance with institutional guidelines.
Experimental animals
In this study, 69 male Sprague-Dawley rats aged between 9 and 10 weeks (280–320 g) were housed in a temperature-controlled environment with a 12:12 h light/dark cycle. In cages, 3–4 rats had free access to water and feed. Animals were closely observed for at least one week before the studies.
LPS-induced sepsis model
To generate the LPS-induced sepsis rat model, we intraperitoneally injected 1.5 mg/kg LPS (from Salmonella typhosa; L6386, Sigma-Aldrich, USA) in 1 ml physiological saline into the rat abdomen. When required, animals were anesthetized by the intraperitoneal injection of ketamine (60 mg/kg) and xylazine (10 mg/kg).
RIPoC
RIPoC was induced by applying a pneumatic cuff to the thigh, inflating it to 300 mmHg, and performing three 5 min cycles of limb ischemia followed by 5 min reperfusion over 30 min [13]. The absence of a femoral pulse and purple foot discoloration indicated effective bloodstream occlusion.
Harvesting
After investigations, 10 ml blood was taken via direct intracardiac puncture using 10 ml disposable syringes and 21 gauge needles. For biochemical analyses, plasma was separated from blood by centrifugation at 3000 rpm for 15 min. Liver tissue from the left lobe was prepared after laparotomy under anesthesia. The liver was sectioned into squares from the center and divided for experiments.
Experimental protocols and experimental groups
Protocol 1. LPS-induced hepatic injury
To investigate the effects of LPS-induced sepsis on the liver, the rats (n = 15) were divided into five groups of three animals each and intraperitoneally administered 1.5 mg/kg LPS (Fig. 1A). Blood and liver tissue were collected from the groups at 0 h (control group), 2 h, 6 h, 12 h, and 18 h after LPS injection.
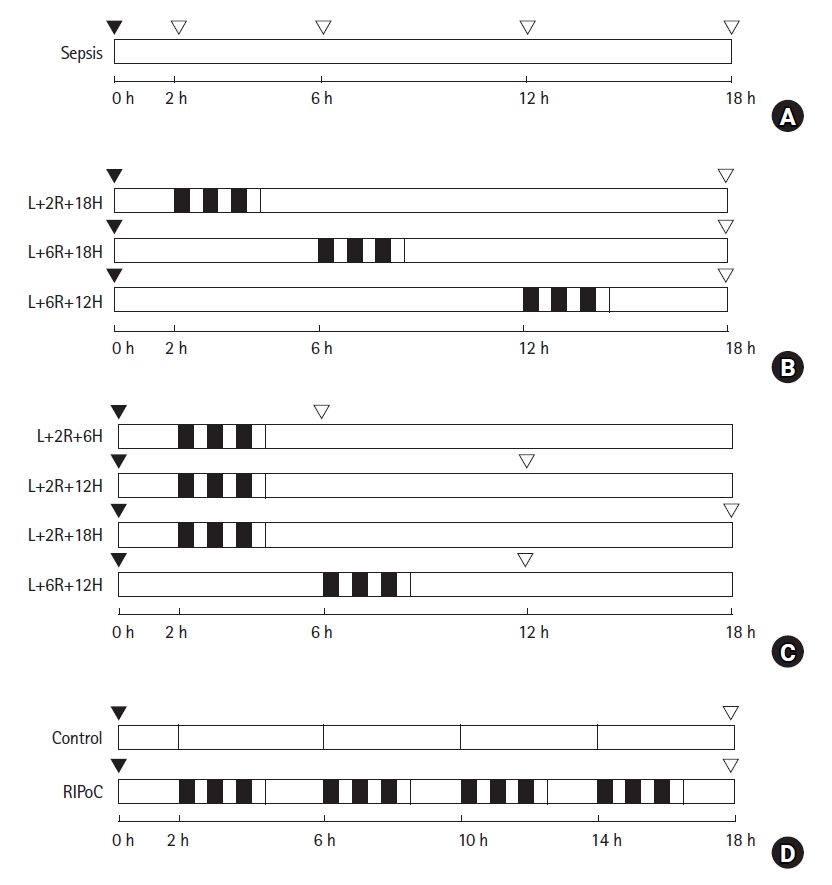
Schematic overview of the study design and experimental groups. (A) To investigate the effects of LPS-induced sepsis on the liver, rats (n = 15) were intraperitoneally administered 1.5 mg/kg LPS. Blood and liver tissue samples were collected after 0, 2, 6, 12, and 18 h. (B) To investigate the effect of RIPoC on LPS-induced hepatic injury, rats (n = 18) were randomly assigned to three groups. (C) To examine duration of RIPoC effects, rats (n = 24) were randomly assigned to four sampling groups (n = 6). (D) To examine repeated RIPoC effects, rats (n = 12) were divided into control and RIPoC groups. Both groups were injected intraperitoneally with ketamine (60 mg/kg) and xylazine (10 mg/kg) to induce anesthesia.  RIPoC: Three cycles (Rt. hind limb ischemia for 5 min + reperfusion for 5 min), ▼ LPS intraperitoneally injected, ▽ sample, L+2R+18H: RIPoC performed at 2 h and sampling at 18 h after LPS injection, L+6R+18H: RIPoC performed at 6 h and sampling at 18 h after LPS injection, L+12R+18H: RIPoC performed at 12 h and blood and liver tissue sampling at 18 h after LPS injection, L+2R+6H: RIPoC performed at 2 h and blood and liver tissue sampling at 6 h after LPS injection, L+2R+12H: RIPoC performed at 2 h and blood and liver tissue sampling at 12 h after LPS injection, L+6R+12H: RIPoC performed at 6 h and blood and liver tissue sampling at 12 h after LPS injection. LPS: lipopolysaccharide, RIPoC: remote ischemic postconditioning.
RIPoC: Three cycles (Rt. hind limb ischemia for 5 min + reperfusion for 5 min), ▼ LPS intraperitoneally injected, ▽ sample, L+2R+18H: RIPoC performed at 2 h and sampling at 18 h after LPS injection, L+6R+18H: RIPoC performed at 6 h and sampling at 18 h after LPS injection, L+12R+18H: RIPoC performed at 12 h and blood and liver tissue sampling at 18 h after LPS injection, L+2R+6H: RIPoC performed at 2 h and blood and liver tissue sampling at 6 h after LPS injection, L+2R+12H: RIPoC performed at 2 h and blood and liver tissue sampling at 12 h after LPS injection, L+6R+12H: RIPoC performed at 6 h and blood and liver tissue sampling at 12 h after LPS injection. LPS: lipopolysaccharide, RIPoC: remote ischemic postconditioning.
Protocol 2. The effects of RIPoC on LPS-induced hepatic injury
The rats (n = 18) were randomly assigned to three groups (n = 6/group). After intraperitoneal injection with LPS, RIPoC was performed at 2 h, 6 h, and 12 h and blood and liver tissue taken at 18 h (Fig. 1B).
Protocol 3. Assessing RIPoC duration
After the intraperitoneal injection of LPS, the rats (n = 24) were randomly assigned to the following four groups (n = 6): (1) RIPoC performed at 2 h and blood and liver tissue sampling at 6 h after LPS injection (L+2R+6H group); (2) RIPoC performed at 2 h and sampling at 12 h after LPS injection (L+2R+12H group); (3) RIPoC performed at 2 h and sampling at 18 h after LPS injection (L+2R+18H group); and (4) RIPoC performed at 6 h and sampling at 12 h after LPS injection (L+6R+12H group) (Fig. 1C).
Protocol 4. The effects of repeated RIPoC
To confirm the effects of repeated RIPoC (performed at four times at regular 4 h intervals), the rats (n = 12) were intraperitoneally injected with LPS. Then, RIPoC was alternately performed on the right and left legs at 2 h, 6 h, 10 h, and 14 h (four times in total). To exclude the effects of ketamine on the liver (administered intraperitoneally for sedation and analgesia during RIPoC), a control group received only ketamine at 2 h, 6 h, 10 h, and 14 h (Fig. 1D). At 18 h, blood and liver tissue were collected from all the rats in both groups.
Serum liver enzyme measurements
Serum alanine transaminase (ALT) and aspartate transaminase (AST) levels were measured spectrophotometrically using a Vitros 250® instrument (Johnson & Johnson, USA) according to manufacturer’s instructions.
Hepatic tissue malondialdehyde (MDA) levels and superoxide dismutase (SOD) activity
MDA levels that indicated lipid peroxidation were measured spectrophotometrically (Smartspec 3000 spectrophotometer, Bio-RAD, USA) using thiobarbituric acid reactive substance assays [14]. Hepatic tissue was homogenized (Kontes Glass Co., USA) and 15% trichloroacetic acid (Sigma-Aldrich), 0.375% thiobarbituric acid (Alfa Aesar, USA), and 0.25 N HCl were added to 0.1 ml of the homogenate. The solution was then boiled for 15 min before cooling to room temperature. Then, after 10 min of centrifugation at 12,000 rpm, absorbance values were measured spectrophotometrically at 535 nm. Protein concentrations were determined using the Bradford assay and MDA activity expressed as ‘nmol/mg’ protein.
Superoxide dismutase (SOD) activity assays were performed using the pyrogallol method [15]. Tris-HCl (50 mM) and 1 mM pentetic acid buffer were used as reaction media, and pyrogallol (20 mM, 10 ml) in 10 mM HCl buffer and 10 ml 0.1 methylenediaminetetraacetic acid buffer were added. Homogenized hepatic tissue was then added to the reaction mixture and decreased pyrogallol absorbance values were monitored spectrophotometrically (Smartspec 3000 spectrophotometer, Bio-RAD, USA) at 420 nm. SOD activity was determined as the amount of enzyme that reduced the color change by 50% (i.e., 50% inhibition of pyrogallol auto-oxidation) and expressed as ‘U/mg’ protein [16].
Western blotting
Hepatic tissue was homogenized in radioimmunoprecipitation assay buffer (Thermo Fisher Scientific, USA) before centrifugation for 20 min at 12,500 rpm and 4°C. Supernatants were then protein extracted. Blotting membranes were blocked in 1% bovine serum albumin in Tris-buffered saline plus 0.1% Tween 20 for 2 h at room temperature. After thorough washing, blots were incubated overnight at 4°C with antibodies against tumor necrosis factor-α (TNF-α), nuclear factor-κB (NF-κB) (1 : 1,000; Santa Cruz Biotechnology, USA), and β-actin (1 : 5000; Cell Signaling Technology, USA). The activated form of NF-κB is a heterodimer of p50/52 and p65. In this study, quantification was performed using a p65 antibody.
Washed membranes were then incubated with secondary anti-mouse horseradish peroxidase-conjugated antibodies (1 : 1,000; Cell Signaling Technology, USA) and washed. Protein bands were reacted with an enhanced chemiluminescence substrate kit (Advansta, USA) and visualized using medical radiography film [17]. Image J software (ver. 1.47; National Institutes of Health, USA) was used to quantify protein signal intensity.
Histopathological analyses
Liver specimens were promptly fixed in 10% buffered formalin for 24 h at room temperature before embedding in paraffin, cutting into 5 μm sections, and staining with hematoxylin and eosin (HE). Four liver tissue samples from each group were used, and 16 random high-power fields/slides were blindly analyzed for polymorphonuclear neutrophil (PMN) counts under light microscopy at 400× in a blinded fashion. Average values were reported.
Statistical analyses
Data were expressed as the mean ± standard error of the mean. For analyses, a one-way analysis of variance was performed, followed by Tukey HSD (Honestly Significant Difference) tests for post hoc comparisons. Differences between the groups were considered significant at P < 0.05. Data analyses were performed using SPSS® software version 23.0 for Windows (SPSS Inc., USA).
Results
In protocol 1 analyses, serum ALT and AST levels were higher in the L+2H, L+6H, L+12H, and L+18H groups when compared with the control group (P < 0.001), but no significant differences were identified between the L+12H and L+18H groups (Fig. 2A). For MDA and SOD measurements that reflected cellular lipid peroxidation caused by oxidative stress no significant changes in the L+2H group were observed when compared with the control group, whereas in the L+6H, L+12H, and L+18H groups, MDA was significantly higher and SOD lower when compared with the control group (P < 0.05). When compared with the L+12H group, similar results were observed in the L+18H group (Figs. 2B and C). TNF-α and NF-κB levels were higher in the L+2H, L+6H, L+12H, and L+18H groups when compared with the control group (P < 0.05) but they plateaued after 6 h (Figs. 2D and E).
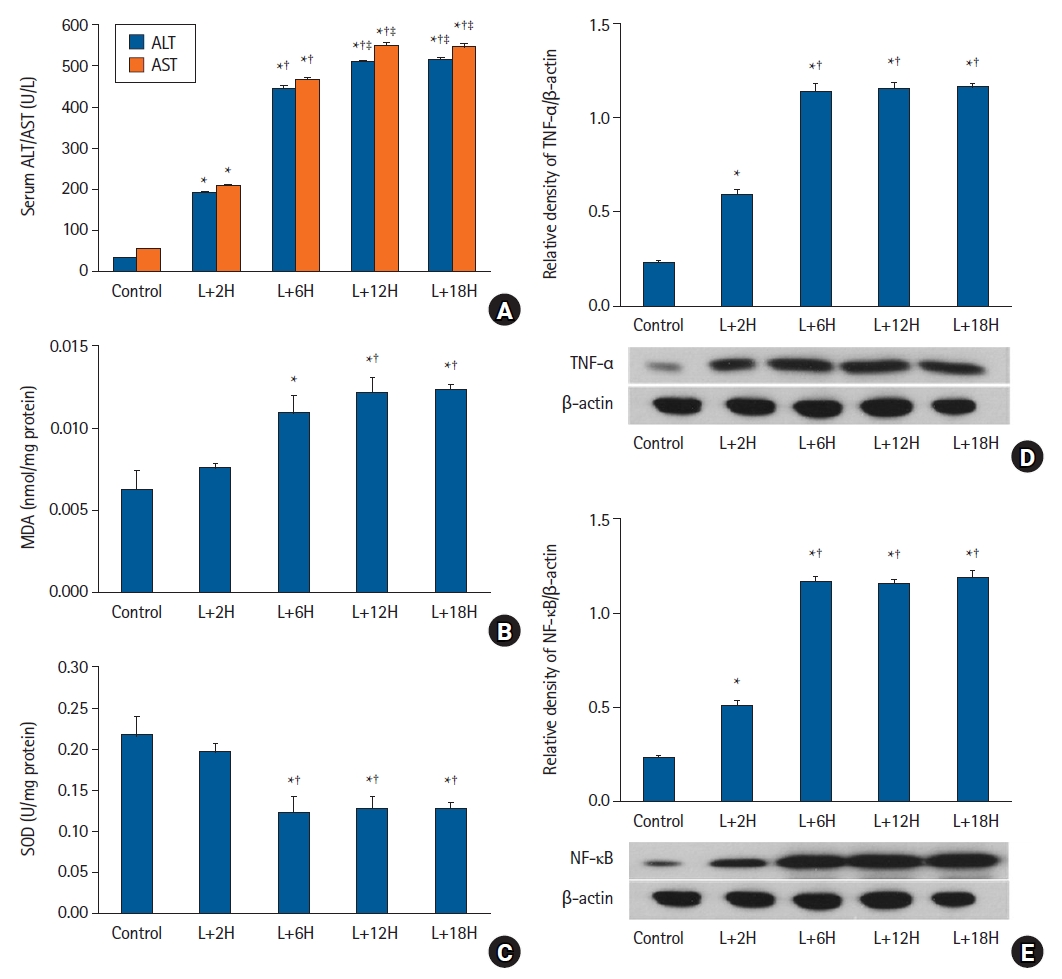
The LPS-induced sepsis model. (A) Serum ALT and AST levels increased over time (P < 0.001) but plateaued after approximately 12 h. (B) Hepatic tissue MDA levels did not significantly change until 2 h and increased up to 12 h. (C) Hepatic tissue SOD activity did not significantly change until 2 h and decreased up to 12 h. (D) TNF-α and (E) NF-κB expression levels showed similar patterns to MDA. *P < 0.05 versus the sham group, †P < 0.05 versus the L+2H group, ‡P < 0.05 versus the L+6H group. Data are expressed as the mean ± standard error of the mean (n = 3 rats/group). LPS: lipopolysaccharide, ALT: alanine transaminase, AST: aspartate transaminase, MDA: malondialdehyde, SOD: superoxide dismutase, TNF-α: tumor necrosis factor-α, NF-κB: nuclear factor-κB.
In protocol 2 analyses, serum ALT and AST and hepatic tissue MDA levels were lower, whereas SOD activity was higher in the L+6R+18H and L+12R+18H groups when compared with the L+2R+18H group (P < 0.001). ALT and AST levels were significantly different between the L+6R+18H and L+12R+18H groups (P < 0.001) (Fig. 3A); however, no significant differences were recorded for MDA levels and SOD activity (Figs. 3B and C).
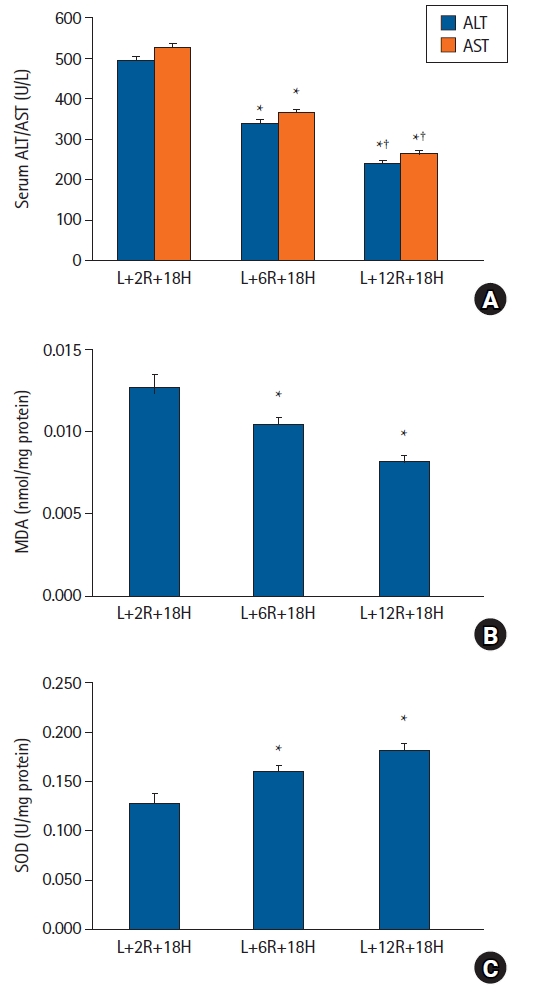
The effects of RIPoC on LPS-induced hepatic injury. (A) Serum ALT and AST levels in L+6R+18H and L+12R+18H groups were lower when compared with the L+2R+18H group, while levels in the L+12H+18H group were also lower when compared with the L+6R+18H group. (B) Hepatic tissue MDA levels in L+6R+18H and L+12R+18H groups were lower when compared with the L+2R+18H group. (C) Hepatic tissue SOD activity in L+6R+18H and L+12R+18H groups were higher when compared with the L+2R+18H group. *P < 0.05 versus the L+2R+18H group, †P < 0.05 versus the L+6R+18H group. Data are expressed as the mean ± standard error of the mean (n = 6 rats/group). RIPoC: remote ischemic postconditioning, LPS: lipopolysaccharide, ALT: alanine transaminase, AST: aspartate transaminase, MDA: malondialdehyde, SOD: superoxide dismutase.
In protocol 3 analyses, serum ALT and AST levels in the L+2R+6H group were significantly lower when compared with the L+2R+12H and L+2R+18H groups (P < 0.001); however, no differences were observed between the L+2R+12H and L+2R+18H groups. In the L+6R+12H group, liver enzyme levels were lower when compared with the L+2R+12H group (P < 0.001) that were sampled at the same time but postconditioned at different times (Fig. 4A). MDA levels were lower and SOD activity was higher in the L+2R+6H group when compared with the L+2R+12H and L+2R+18H groups (P < 0.001). When compared with the L+2R+12H group, MDA levels were low and SOD activity was high in the L+6R+12H group (P < 0.001) (Figs. 4B and C).
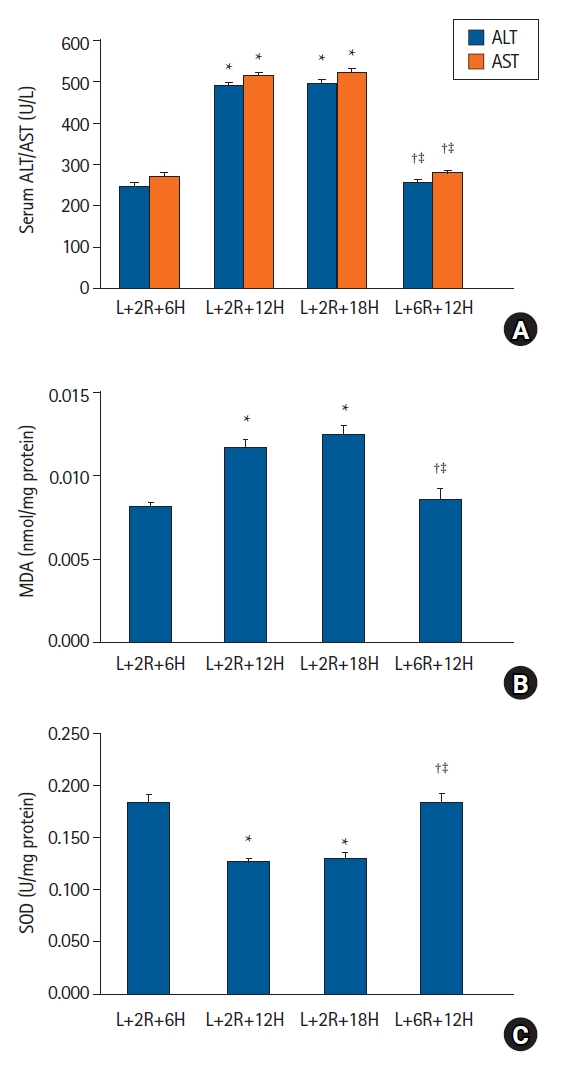
Duration of RIPoC effects. (A) Serum ALT and AST levels in the L+2R+6H group were lower when compared with L+2R+12H and L+2R+18H groups. Levels in the L+6R+12H group were lower when compared with L+2R+12H and L+2R+18H groups. (B) Hepatic tissue MDA levels also showed similar patterns to serum ALT and AST levels. (C) Hepatic tissue SOD activity in the L+2R+6H group was higher when compared with L+2R+12H and L+2R+18H groups. Levels in the L+6R+12H group were higher when compared with L+2R+12H and L+2R+18H groups. *P < 0.05 versus the L+2R+6H group, †P < 0.05 versus the L+2R+12H group, ‡P < 0.05 versus L+12R+18H group. Data are expressed as the mean ± standard error of the mean (n = 6 rats/group). RIPoC: remote ischemic postconditioning, ALT: alanine transaminase, AST: aspartate transaminase, MDA: malondialdehyde, SOD: superoxide dismutase.
In protocol 4 analyses, serum ALT, AST, and MDA levels were lower while SOD activity was higher in the RIPoC group (subjected to RIPoC four times at regular intervals) when compared with the control group (P < 0.05) (Figs. 5A-C). Furthermore, TNF-α and NF-κB levels in the RIPoC group (western blotting) were lower when compared with the control group (P < 0.05) (Figs. 5D and E). In the HE-analyzed liver tissue, the degree of PMN infiltration in the RIPoC group was lower when compared with the control group (Fig. 6A). Intrahepatic sinusoidal neutrophil numbers were significantly decreased in the RIPoC group when compared with the control group numbers (Fig. 6B).
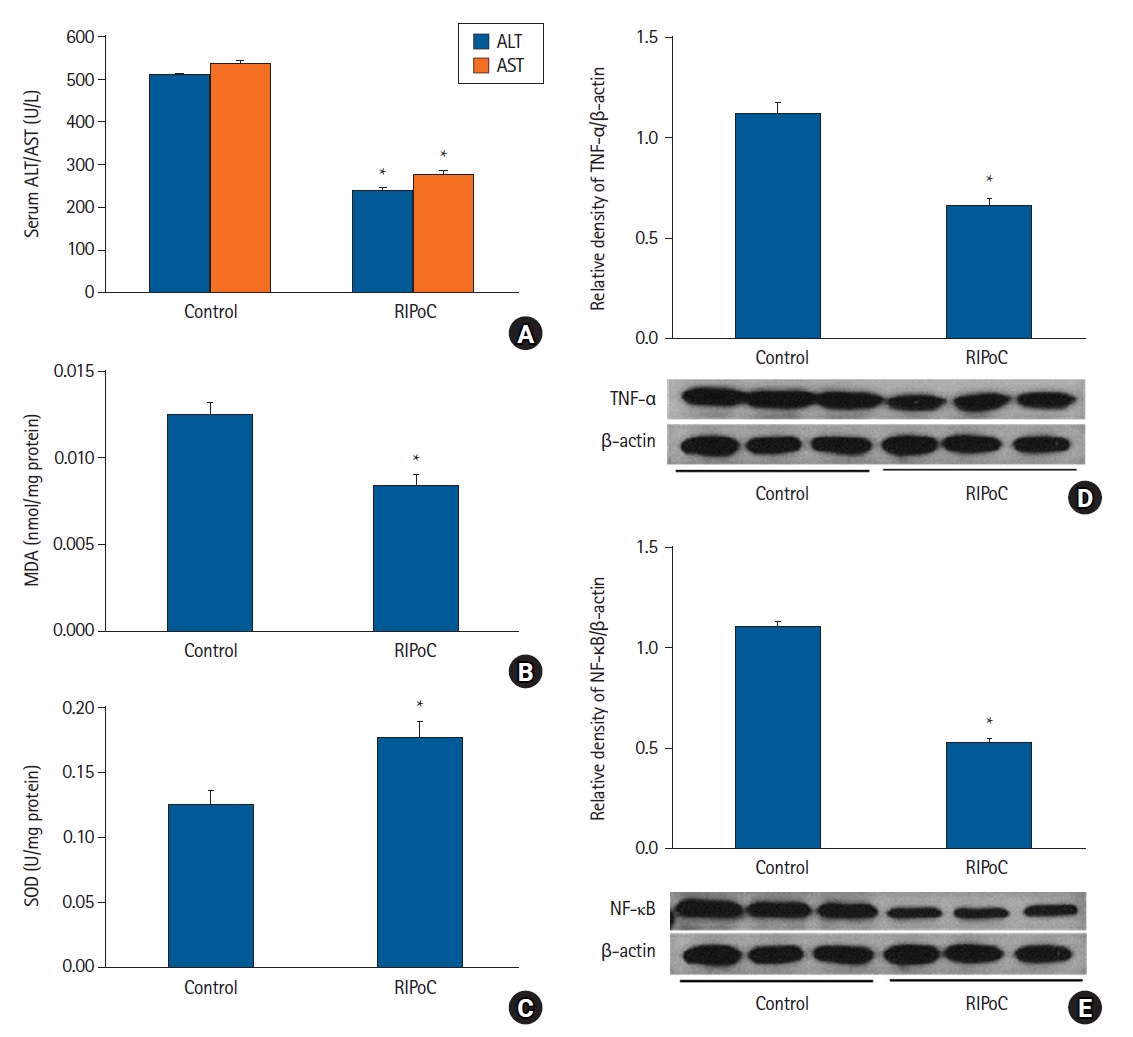
Repeated RIPoC effects. (A) Serum ALT and AST levels in the RIPoC group were significantly lower when compared with the control group. (B) Hepatic tissue MDA levels in the RIPoC group showed similar patterns to ALT and AST levels. (C) Hepatic tissue SOD activity in the RIPoC group was maintained when compared with the control group. (D) TNF-α and (E) NF-κB expression levels were also significantly lower when compared with the control group. *P < 0.05 versus the control group. Data were expressed as the mean ± standard error of the mean (n = 6 rats/group). RIPoC: remote ischemic postconditioning, ALT: alanine transaminase, AST: aspartate transaminase, MDA: malondialdehyde, SOD: superoxide dismutase, TNF-α: tumor necrosis factor-α, NF-κB: nuclear factor-κB.
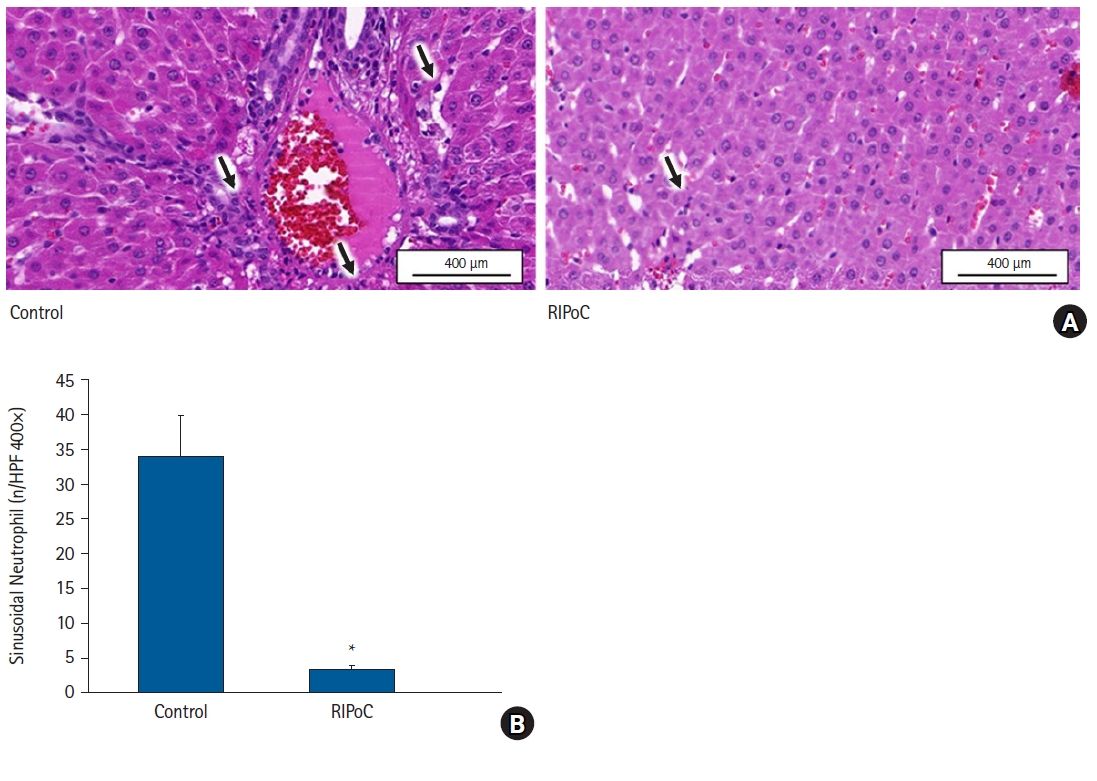
Histopathological analyses. (A) PMN infiltration in liver sinusoid and perivascular areas was significantly reduced in the RIPoC group when compared with controls (arrows) (400×). (B) Intrahepatic sinusoidal neutrophil numbers in 16 random high-power fields (400×). *P < 0.05 versus the control group. Data are expressed as the mean ± standard error of the mean (n = 6 rats/group). PMNs: polymorphonuclear neutrophils, RIPoC: remote ischemic postconditioning.
Discussion
We showed that RIPoC stimulus via transient hind limb ischemia significantly alleviated LPS-induced liver injury in a septic rat model. In all the RIPoC treatment groups, serum ALT and AST levels were reduced, indicating that RIPoC protected cells against liver injury. RIPoC appeared to function as an immunological and antioxidant modulator in sepsis based on significant reductions in inflammatory and oxidative stress markers after its application [18].
RIC is an interesting and clinically significant tissue conditioning modality. The techniques is minimally invasive and may be applied to transplantation, vascular surgery, and cardiac surgery settings [19]. Brief I/R cycles at distant tissue may protect organs when subjected to potentially lethal ischemia [20,21]. Short IR cycles in distant tissue release humoral factors such as interleukins (IL), adenosine, opioids, bradykinin, and nitric oxide (NO) [22]. Under ischemia, these humoral factors and direct stimuli are detected by remote organ innervation [23]. These protective signals may activate G-protein-coupled receptors or receptor tyrosine kinases that induce the reperfusion injury salvage kinase (RISK) pathway, or inflammatory cytokines via glycoprotein 130 or the TNF receptor to activate the survivor activating factor enhancement (SAFE) pathway [24]. Although the exact molecular mechanisms underpinning these protective actions remains unclear, accumulating evidence now suggests that they suppress inflammatory TNF-α and NF-κB gene expression [25], decrease reactive oxygen species (ROS) production, and suppress human neutrophil adhesion and phagocytosis. Such altered immune profile have facilitated their investigation and application in sepsis. Honda et al. [8] reported that RIC improved cardiac output and survival in a LPS-induced sepsis model. Kim et al. [26] also observed that RIPoC alleviated inflammatory responses and increased survival rates by modifying NF-κB-mediated cytokine expression in a LPS-induced sepsis model. Previous studies also reported that using RIPC or RIPoC strategies in a sepsis setting reduced inflammatory response while increasing survival; however, it is unclear if RIPC or RIPoC increased hepatic cell anti-oxidative defenses. Our data suggested that RIPoC improved hepatic function by increasing not only anti-inflammatory outputs in hepatic cells but also antioxidant defenses.
The liver is the primary site for bacterial clearance; it induces inflammatory responses to sepsis by producing acute phase proteins and proinflammatory cytokines [6,7]. NF-κB is a key transcription factor that regulates proinflammatory cytokine genes (e.g., TNF-α, IL-1β, and IL-6), adhesion molecules, chemokines, and monocyte chemotactic protein-1 [27]. Therefore, NF-κB is activated early in response to sepsis in injured organs [26]. The NF-κB pathway blockade remediates the pathological abnormalities induced by sepsis in animal models while inhibited NF-κB activation reduces multiple inflammatory gene expression, preventing multiple organ damage and increasing survival rates after sepsis. Therefore, effective NF-κB inhibition or reduced TNF-α levels are significant therapeutic objectives in sepsis treatment [28]. In the sepsis models, higher NF-κB and TNF-α levels are associated with higher mortality rates and poor clinical outcomes. Our data indicate a marked elevation in the NF-κB and TNF-α levels after LPS intraperitoneal injection, while a marked reduction in TNF-α levels was observed after RIPoC.
In sepsis, bacteria and endotoxins directly act on phagocytes that cause lipid peroxidation in cellular membranes and severe damage to cellular structure and function [29]. Free radical production and oxidative stress in ischemia cause tissue damage that is accompanied by activated inflammatory responses. Previous research reported that oxidative stress is one of the key mechanisms implicated in hepatic ischemia damage processes [30]. MDA and SOD are two major pathophysiological markers used to assess free radical metabolism [31]. MDA is an oxidative stress biomarker induced by lipid peroxidation and is the end-product of polyunsaturated fatty acid [32]. MDA levels also reflect the degree of cell injury from the reactive oxygen metabolites [33]. SOD is a key endogenous antioxidant enzyme that scavenges ROS and helps maintain mitochondrial functional integrity [17]. In our study, SOD activity in hepatic tissue was significantly decreased whereas MDA levels were significantly increased in LPS-induced sepsis rats when compared with controls. Thus, RIPoC satisfactorily reduced oxidative stress. Furthermore, the anti-inflammatory and anti-oxidative stress effects of RIPoC were putatively correlated with liver protection. The simultaneous reduction in NF-κB and TNF-α levels, and also improved oxidative stress profiles after RIPoC administration, suggested complex pathway interplay leading to liver protection. Further molecular and genetic level studies are required to fully understand this mechanistic complexity.
RIC, with different insult timing, has been used in different clinical settings. Some studies have used preconditioning that induces ischemic preconditioning prior to the ischemic insult. In our study, RIC was performed after sepsis, the RIPoC approach. In clinical settings, preconditioning before septic insult is often unfeasible, although both types of ischemic conditioning may be beneficial. To establish optimal RIPoC timing, RIPoC was performed at three different time points (2 h, 6 h, and 12 h after insult), and inflammatory and oxidative stress indicators assessed at 18 h post insult. The 12 h time point was associated with greater liver protection when compared with the 6 h time point. Also, to verify if improved anti-inflammatory and antioxidant defenses sustained responses, anti-inflammatory and antioxidant parameters were measured at 6 h, 12 h, and 18 h after the insult after performing RIPoC at 2 h after insult. When RIPoC was conducted at 2 h after insult, no improvements were observed at 12 and 18 h, indicating that RIPoC only temporarily improved liver anti-inflammatory and antioxidant parameters. This finding was consistent with Costa et al. [22] who demonstrated that RIC functioned via the temporary and short-term enhancement of liver and kidney cell antioxidant defenses to avoid the deleterious consequences of future IR injury. These observations are important for the clinical translation of RIPoC because RIPoC-induced protection may be transitory and limited-duration events. Therefore, the effects of repeated RIPoC episodes on molecular outcomes were also examined. Reports on the effects of repeated RIPoC have increased in recent years. Wei et al. [34] were the first to report that performing RIC daily for a month after myocardial infarction decreased peri-infarct inflammatory responses and enhanced cardiac functional recovery and survival. Honda et al. [8] reported that repeated RIC exerted additional benefits with respect to mortality during sepsis. Consistent with previous studies, the rats in our study subjected to repeated RIPoC demonstrated improved hepatic function when compared with control rats. Furthermore, repeated RIPoC improved anti-inflammatory and antioxidant profiles. Histological liver section examinations from LPS-induced sepsis rats revealed infiltrated neutrophils, vacuolization, and necrotic hepatocytes, while these histological changes were attenuated by repeated RIPoC. However, the mechanisms underpinning repeated RIPoC applications have not been fully elucidated in previous reports and in our study; therefore, future studies are required to determine the precise mechanisms.
Our study had some limitations. For example, increasing anti-inflammatory and antioxidant capacity levels are not the only effects expected upon RIC. RISK and SAFE pathway activation, NO synthase activity, and NO release are variables that change with RIC, but they were not analyzed in our study. Moreover, survival rates were also not assessed in this study. Nevertheless, we provided a scientific foundation and a viable functional model for RIPoC-induced hepatic protection during sepsis, implying that RIPoC could be a potential therapeutic approach for sepsis treatment.
In conclusion, we comprehensively demonstrated that RIPoC attenuated liver injury in an LPS-induced sepsis model by modifying inflammatory responses and oxidative stress response for a limited period. Although additional repeated RIPoC improved hepatic injury during sepsis, further studies are required to fully understand the additional benefits and specific molecular mechanisms underpinning repeated RIPoC.
Notes
Funding
None.
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.
Data Availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author Contributions
Jin Duck Cho (Conceptualization)
Hoon Jung (Conceptualization)
Jeong Eun Lee (Data curation)
Eun Kyung Choi (Conceptualization)
Hyun Ah Kim (Data curation)
Hyun-Su Ri (Conceptualization)
Hyunjee Kim (Conceptualization)
Ji Young Park (Methodology)
Kyung-Hwa Kwak (Conceptualization)
Dong Gun Lim (Conceptualization; Supervision)