Introduction
Thiamylal sodium is a short-acting barbiturate used primarily for anesthesia induction and also in the management of intracranial hypertension [1]. Thiamylal-induced arterial vasodilation and myocardial depression are generally relatively mild [2]. Barbiturates are used in alkaline solutions (pH 10.5ŌĆō11.5) in many clinical settings, and following their venous injection, have been reported to cause phlebitis or thrombophlebitis [3]. However, there is no previously reported study on the effects of barbiturates on the inflammation response, or injury and proliferation of vascular smooth muscle cells (VSMCs).
In this study, we first screened clinical-grade thiamylal so dium solution (TSS: Citosol)-responsive inflammatory related genes in a rat VSMC cell line using microarray analysis. Endogenous proinflammatory cytokines, including interleukin (IL)-1╬▒, have been reported to promote a proliferative and proinflammatory phenotype in human VSMCs [4]. Inflammation and the proliferation of VSMCs, as well as cytokine secretion after vascular injury, contribute to the pathogenesis of atherosclerosis or in-stent restenosis, which can result in vessel bypass graft failure [5,6]. After an initial injury, growth factors and cytokines promote VSMC proliferation, migration, and extracellular matrix protein deposition [7]. We thus examined whether TSS could increase proinflammatory cytokine secretion from VSMCs and promote cell proliferation.
Materials and Methods
Materials
Dulbecco's modified Eagle's medium (DMEM) was purchased from Gibco BRL (Grand Island, NY, USA). Fetal bovine serum (FBS) was purchased from JRH Biosciences Inc. (Lenexa, KS, USA). Bovine serum albumin (BSA) was purchased from Sigma Chemical Co. (St. Louis, MO, USA). TSS (Citosol) was purchased from Kyorin Pharmaceutical Co. (Tokyo, Japan). Enzyme-linked immunosorbent assay (ELISA) kits for rat IL-1╬▒ and IL-6 were purchased from R&D Systems, Inc. (Minneapolis, MN, USA) and BioSource International (Camarillo, CA, USA), respectively. The Water Soluble Tetrazolium-1 (WST) Cell Proliferation Assay System was purchased from Takara Bio (Shiga, Japan). All other chemical reagents, including sodium carbonate, were purchased from Wako Pure Chemicals (Osaka, Japan) unless otherwise stated.
Cell culture
Rat VSMCs (American Type Culture Collection, USA) were maintained in low-glucose DMEM supplemented with 5% FBS at 37Ōäā in a humidified atmosphere of 95% air and 5% CO2. VSMCs were grown to confluence, then cultured for an additional 2 days in DMEM with 0.1% BSA prior to experimental use.
Microarray analysis
Rat VSMCs were treated with TSS for 3 h and total RNA was prepared using the acid guanidinium thiocyanate-phenolchloroform (AGPC) extraction method. Affymetrix GeneChip (Santa Clara, CA, USA) expression analysis, including sample preparation, hybridization and data analysis, was performed as described previously [8].
Quantitative reverse transcription polymerase chain reaction (qRT-PCR)
Total RNA samples from VSMCs treated with different concentrations of TSS or sodium carbonate were prepared using the AGPC extraction method with ISOGEN (Nippon Gene, Tokyo, Japan) according to the manufacturer's instructions. Total RNA (0.4 ┬Ąg) was reverse-transcribed using Moloney murine leukemia virus RT (ReverTra Ace-╬▒ kit; Toyobo, Osaka, Japan). IL and toll-like receptor (TLR) mRNA levels were measured using the SYBR-green (Toyobo) RT-PCR method and normalized to hypoxanthine phosphoribosyltransferase 1 (Hprt1) expression (n = 4) [9]. The primer sequences used in this study are listed in Table 1. An ABI Prism 7500 Sequence Detection System (Applied Biosystems, Foster City, CA, USA) was used for detecting gene expression. Values are expressed as a percent of control culture (100%).
Cell proliferation assay
VSMCs were plated in 96-well plates in DMEM. The culture medium was replaced the following day with fresh medium containing different concentrations of TSS and the VSMCs were cultured for a further 24 h. Proliferation was measured using the WST-1 assay (n = 10) [10]. Briefly, premixed WST-1 reagent (10 ┬Ąl) was added to each well and VSMCs were incubated for 4 h. Sample absorbance, compared with a background blank control, was measured at 450 nm using a microplate reader according to the manufacturer's instructions.
Quantification of rat IL-6 by ELISA
Based on our previous preliminary data to detect a significant increase in IL-6 protein level [11], VSMCs were stimulated with TSS for 24 h. VSMC culture supernatants were collected and centrifuged (12000 rpm, 1 min). Supernatants were stored at ŌĆō80Ōäā until later use in assays. ELISA was carried out to determine IL-6 protein expression levels according to the manufacturer's instructions (n = 8).
Ex vivo stimulation of rat aorta
Nine-week-old Sprague Dawley rats were purchased from Kyudo Co. (Saga, Japan). Animal handling was supervised by the Institutional Animal Experiment Committee. Rats were euthanized under deep anesthesia with pentobarbital and ketamine. The thoracic-aorta was excised and the adventitia was removed. The aorta was cut into small pieces and about 5 g (wet weight) of tissue was stimulated with or without 30 ┬ĄM TSS in 500 ┬Ąl DMEM supplemented with 0.1% BSA for 24 h in 12-well plates [11]. Supernatants were collected and centrifuged (12,000 rpm, 1 min) prior to being stored at ŌĆō80Ōäā until further use in assays to measure IL-1╬▒ or IL-6 production by ELISA (n = 5). The IL-6 concentration in the supernatant was normalized with the wet weight of the aortic segment.
Measurement of cell viability
Confluent VSMCs were serum-starved for 24 h prior to treatment with TSS. After 24 h incubation, attached cells were harvested with trypsin-EDTA. Cells in the medium were collected by centrifugation and then stained with 0.4% trypan blue. Numbers of total and dead cells were counted using a hemocytometer (n = 8) [12]. Values are expressed as a percent of control culture (100%).
Statistical analysis
Normality of the data was assessed with the Shapiro-Wilk test. A t-test was used to compare between two groups. Evaluation of differences between multiple groups was performed using one-way analysis of variance (ANOVA) and Fisher's post hoc test, as appropriate. All analyses were performed using JMP Pro software (ver. 11; SAS Institute Inc., Cary, NC, USA). Data are presented as means ┬▒ standard error of the mean (SEM). A probability value of P < 0.05 with a 95% confidence interval was considered to indicate a statistically significant difference.
Results
Microarray analysis
Following TSS-responsive gene screening in rat VSMCs using a microarray, TSS was found to enhance expression of several genes involved in inflammation (Table 2).
TSS enhances inflammatory response in VSMCs
Exposure of VMSCs to TSS at a dose of 100 ┬ĄM resulted in upregulation of IL-1╬▒ mRNA (3,262.9% ┬▒ 1,195.0% vs. Control (100%), P = 0.005; Fig. 1A). However, TSS at doses of 3, 10, 30, and 100 ┬ĄM resulted in downregulation of IL-1╬▓ mRNA (15.9% ┬▒ 4.1% vs. Control, P = 0.045; 4.2% ┬▒ 4.1%, P = 0.022; 17.2% ┬▒ 9.2%, P = 0.049; 4.5% ┬▒ 2.8%, P = 0.023, respectively; Fig 1B) in a dose-dependent manner (Figs. 1A and 1B). A similar trend was observed for IL-6 mRNA (443.1% ┬▒ 61.2% vs. Control, P < 0.001 at 100 ┬ĄM; Fig. 1C).
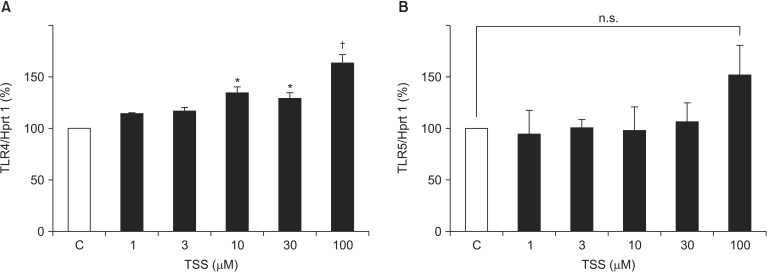
Receptor-mediated activation of VSMCs can contribute to increased inflammatory responses; thus, we next investigated TLR expression patterns. VSMCs stimulated by TSS at doses of 30 and 100 ┬ĄM showed a mild but significant increase in TLR4 mRNA expression in a dose-dependent manner (134.5% ┬▒ 5.7% vs. Control, P = 0.001; 163.5% ┬▒ 7.9%, P < 0.001, respectively; Fig. 2A). Although TLR5 mRNA showed a tendency to increase, the difference was not significant (P = 0.152 at 100 ┬ĄM; Fig. 2B). Finally, levels of TLR2, TLR3, and TLR6 mRNA expression were very low and did not change markedly.
TSs increases IL-6 production In Vitro and Ex Vivo
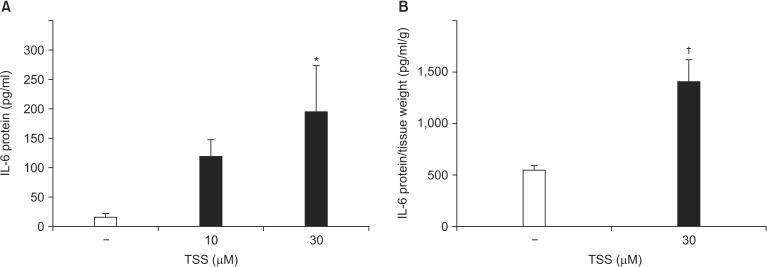
VSMCs were incubated with TSS for 24 h. TSS-induced IL-6 production was significantly increased in a dose-dependent manner (Control 16.4 ┬▒ 6.5 pg/ml [range: 0.91ŌĆō31.85 pg/ml]; TSS [10 ┬ĄM] 119.2 ┬▒ 28.4 pg/ml [range: 51.9ŌĆō186.4 pg/ml], P = 0.145; TSS [30 ┬ĄM] 194.8 ┬▒ 78.1 pg/ml [range: 10.1ŌĆō379.6 pg/ml], P = 0.015; Fig. 3A). To confirm that proinflammatory cytokines were produced from blood vessels, a segment of rat aorta without adventitia was stimulated ex vivo with TSS for 24 h. IL-6 production from rat aorta was increased by TSS stimulation (Control 551.2 ┬▒ 43.5 pg/ml/g [range: 430.3ŌĆō802.4 pg/ml] vs. TSS 1,405 ┬▒ 43.5 pg/ml/g [range: 802.4ŌĆō2,009.0 pg/ml], P = 0.005; Fig. 3B). IL-1╬▒ production in the medium was undetectable under these conditions (data not shown). These ex vivo results are consistent with those obtained during the in vitro experiments.
TSS attenuates cell proliferation and viability
TLR-activated VSMCs have been reported to produce IL-1╬▒ in an autocrine manner, which contributes to proliferation and chemokine release by VSMCs [4]. VSMCs were cultured with TSS for 24 h and the effect of TSS on proliferation was determined. The WST-1 cell proliferation assay showed that a low concentration of TSS increased cell proliferation of VSMCs (percentage proliferation at 10 ┬ĄM TSS: 124% ┬▒ 21% vs. Control [range: 109%ŌĆō140%], P = 0.002; Fig. 4A). In contrast, the highest concentration of TSS inhibited VMSC proliferation (percent proliferation 100 ┬ĄM TSS: 43% ┬▒ 2% vs. Control [range: 40%ŌĆō 46%], P < 0.001).
Cell viability was assessed by trypan blue staining. TSS at the highest concentration significantly reduced the viability of VSMCs (percentage of viable cells at 30 ┬ĄM TSS: 87% ┬▒ 3% [range: 80%ŌĆō94%], P = 0.015; 100 ┬ĄM TSS: 69% ┬▒ 4% [range: 61%ŌĆō77%], P < 0.001; Fig. 4B). It has been reported that VSMCs produce IL-1 receptor antagonist (IL-1Ra) that inhibits the activities of IL-1╬▒ and IL-1╬▓ and regulates both inflammatory responses and cell proliferation [13,14,15]. However, IL-Ra mRNA was not significantly increased by TSS stimulation (100 ┬ĄM TSS: P = 0.521; Fig. 4C).
Effects of sodium carbonate on VSMCs
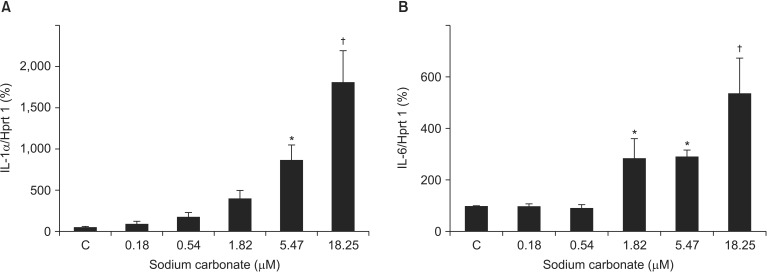
Finally, we investigated the effects of sodium carbonate on VSMCs. TSS contains 0.035 g of sodium carbonate per 0.5 g of thiamylal sodium (0.18 ┬ĄM of sodium carbonate per 1 ┬ĄM of thiamylal sodium). VSMCs were stimulated with the same amount of sodium carbonate contained in TSS (Fig. 1). IL-1╬▒ mRNA levels (1,734.0% ┬▒ 363.2% vs. Control, P = 0.019 at 5.47 ┬ĄM; 3,633.1% ┬▒ 762.8%, P < 0.001 at 18.25 ┬ĄM) and IL-6 mRNA levels (286.9% ┬▒ 73.8% vs. Control, P = 0.046 at 1.82 ┬ĄM; 291.4% ┬▒ 25.1%, P = 0.042 at 5.47 ┬ĄM; 537.5% ┬▒ 133.7%, P < 0.001 at 18.25 ┬ĄM) were significantly upregulated by sodium carbonate stimulation (Fig. 5). However, the increase in levels of ILs was considerably greater using sodium carbonate alone than with TSS (Fig. 1).
Discussion
In the present study, we demonstrated that TSS increased the expression of IL-1╬▒ and IL-6 in VSMCs. Low concentrations of TSS showed a positive effect on the proliferation of VSMCs. TSS contains a sodium carbonate additive because thiamylal sodium is difficult to dissolve at neutral pH. Because sodium carbonate produces hydroxide ions in aqueous solution, thiamylal is always used in strong alkaline solutions in the clinical setting. Consequently, intravenous injection of barbiturates, including TSS, of ten causes phlebitis or local necrosis at the injection site. In this study, sodium carbonate-induced IL-1╬▒ and IL-6 upregulation was greater than TSS-induced upregulation. These results indicate that TSS-induced proinflammatory cytokine production by VSMCs is likely to be caused by sodium carbonate, whereas pure thiamylal sodium itself has a potent anti-inflammatory effect in VSMCs.
A previous study indicated that endogenous IL-1╬▒ acts as an autocrine growth factor for VSMCs, and its pro-proliferative effects are mediated by signaling via the type-1 IL-1 receptor [4,16]. IL-6 is well-known to induce the expression of C-reactive protein, which is a prognostic marker for atherosclerotic cardiovascular disease [17]. Furthermore, IL-6 stimulates the proliferation of VSMCs [18]. These data suggest that IL-6 from blood vessels could be involved in local inflammatory responses that might accelerate atherogenesis. Furthermore, some clinical studies suggest that inflammatory mediators, including oxidized low-density lipoprotein, IL-6, and tumor necrosis factor-╬▒, are associated with an increase in the relative risk of developing cardiovascular disease, especially ischemic heart disease [19,20,21]. We showed that TSS increased proinflammatory cytokine production from rat VSMCs and aortas. These findings indicate that TSS-induced proinflammatory cytokine production might, at least partially, contribute to the progression of atherosclerosis.
Clarke et al. [22] showed that necrotic VSMCs release IL-1╬▒, which subsequently stimulates the surrounding viable VSMCs to release IL-6 and monocyte chemotactic protein (MCP)-1. In the present study, TSS increased the expression of IL-1╬▒ but not IL-1╬▓ mRNA expression. Additionally, the WST-1 assay and trypan blue viability test suggest that high concentrations of TSS reduced cell viability. Both apoptosis and necrosis can occur at high levels in advanced plaques in conjunction with high serum concentrations of inflammatory factors [23]. Considered together, TSS might contribute to the development of VSMC necrosis and cause plaque build-up.
Members of the TLR family play key roles in the innate immune system and are expressed in a cell type-specific manner [24]. TSS-stimulated VSMCs showed a significant increase in TLR4 mRNA expression level. Previous studies have shown that TLR4 is associated with VSMC proliferation [25,26]. In the present study, TSS-induced VSMC proliferation was observed only at a concentration of 10 ┬ĄM. At this concentration, however, IL levels did not significantly increase. Based on these findings, it is presumed that the effect of TSS on VSMC proliferation is associated primarily with TLR4 upregulation. Earlier studies indicate that TLR4 expression on VSMCs is linked to elevated basal secretion of IL-6 [27]. TSS-induced TLR4 upregulation may play a partial role in the secretion of IL-6. Although TLR3 signaling can evoke proinflammatory and proliferative responses in human VSMCs [28], overall levels of TLR3 mRNA expression were very low in our study.
VSMCs can express an intracellular form of IL-1Ra that regulates inflammatory responses and cell proliferation [13]. A previous study showed that the effect of transforming growth factor (TGF)-╬▓1 on IL-1Ra production could be completely inhibited by an anti-IL-1╬▓ antibody [15]. In our study, neither IL-1Ra nor IL-1╬▓ expression were increased in TSS-stimulated VSMCs. IL-1Ra induction in VSMCs might therefore be dependent on IL-1╬▓ expression.
Clinically relevant plasma concentrations of thiamylal (2.7ŌĆō5.0 mg/kg), 2ŌĆō30 min after bolus intravenous injection, are reported to range between 7.8 and 47.0 ┬ĄM [29]. Furthermore, the initial concentration of thiamylal in clinical use is about 200 ┬ĄM [30]. Thus, the concentrations of thiamylal used in our study represent pharmacologically achievable plasma concentrations. However, because of the numerous limitations of in vitro studies, in vivo studies are needed to confirm whether thiamylal causes the development of blood vessel inflammation.
In summary, we showed that TSS increased IL-1╬▒ and IL-6 production both in vitro and ex vivo, and that low concentrations of TSS increased VSMC proliferation. These effects may promote vascular inflammation and lead to the progression of atherosclerosis.