Introduction
Despite newly developed resuscitation therapies and the use of modern antibiotics, sepsis is still a leading cause of death in critically ill patients [1]. The mortality rate in sepsis patients remains as high as 40-70% [2]. Severe sepsis is characterized by hypoperfusion of major organs, leading to multiple organ failure, shock, and death. Inflammation, a physiological response to infection or injury, plays a key role in both health and disease. Several pro-inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-╬▒), interleukin (IL)-1╬▓, and IL-8 promote systemic inflammation. Cytokine-mediated host defense mechanisms induce significant cell and organ injury and play crucial roles in the pathophysiology of sepsis [3].
IL-1╬▓ is produced from the cleavage of the inactive pro-IL-1╬▓ by stimulated leukocytes and is a fundamental contributor to local and systemic inflammatory responses [4]. IL-1╬▓ is a pro-inflammatory cytokine that functions as a critical regulator of the host defense in response to an infection or injury. However, IL-1╬▓ is extremely toxic when present in excess [5]. Elevated systemic levels of IL-1╬▓ may cause hypotension during septic shock and induce capillary leakage in acute lung injury [6]. IL-1╬▓ is also involved in chronic inflammation associated with arthritis, lung fibrosis and atherosclerosis [7-9]. Therefore, strategies to modulate IL-1╬▓ production in inflammatory diseases are of therapeutic interest. In addition, IL-1╬▓ activates the mitogen-activated protein kinase (MAPK) family including the extracellular signal-regulated kinase 1 and 2 (ERK1/2), c-Jun N-terminal kinase (JNK) and p38 MAPK [10].
Nontoxic molecules that modulate macrophage-mediated inflammatory responses may provide a novel therapeutic strategy for the treatment of sepsis. Numerous studies have been performed to identify the new molecules. The manassantin group of structurally related and functionally unique dineolignans were recently isolated from active root extracts of Saururus cernuus L (Saururaceae), also referred to as "lizard tail" [11]. This fragrant aquatic plant is found in the North America as well as in many parts of Asia (Saururus chinensis) [12-14]. Although their structures and relative stereochemistries were only determined recently, the medicinal value of the plant has long been recognized by Native Americans, early colonists and practitioners of Korean traditional medicine for a range of diseases [14,15]. Manassantin A and B have biological activities, such as anti-inflammatory and anti-plasmodial effects [12,16].
The purpose of this study was to investigate the effect of manassantin B on LPS-induced activation of the pro-inflammatory cytokine IL-1╬▓ and intracellular signaling pathway such as the MAPKs signaling system, and the possibility that manassantin B might be a promising agent for reducing inflammatory response.
Materials and Methods
Materials
Manassantin B was donated from the College of Pharmacy, Yeungnam University, Korea. Escherichia coli 055:B5 endotoxin (lipopolysaccharide, LPS) was purchased from Sigma-Aldrich (St. Louis, MO, USA). Antibodies specific for phosphorylated (P)-ERK, P-JNK and p38 MAPK as well as total (T)-ERK, T-JNK and T-p38 MAPK were supplied by Cell Signaling Technologies (Beverly, MA, USA). Enzyme-linked immunosorbent assay (ELISA) kits for IL-1╬▓ were purchased from R&D Systems (Minneapolis, MN, USA). Dulbecco's modified Eagle's medium (DMEM) high glucose/L-glutamine, fetal bovine serum (FBS) and penicillin/streptomycin was obtained from GIBCO (Gaithersburg, MD, USA). Bicinchoninic acid (BCA) protein assay reagent was supplied by Pierce (Rockford, IL, USA).
Cell culture and treatment
The RAW 264.7 murine macrophage cell line was purchased from the Korea Cell Line Bank (Seoul, Korea). The cells were grown in DMEM supplemented with heat-inactivated 10% FBS, penicillin (100 U/ml) and streptomycin sulfate (100 ┬Ąg/ml) in a 5% CO2 incubator at 37Ōäā. The cells were plated at a density of 2 ├Ś 106 cells/well in a 6-well plate or 5 ├Ś 105 cells/well in 24-well plates. For all experiments, the cells were incubated until they reached 80-90% confluence. The adherent macrophages were washed gently with Ca2+ and Mg2+-free Dulbecco's phosphate buffered saline (DPBS) and cultured in serum-free DMEM for 2 h. Then, the cells were incubated in the absence or presence (1, 3 and 10 ┬ĄM) manassantin B without or with LPS (100 ng/ml). Manassantin B dissolved in phosphate-buffered saline (PBS) was added to the medium 1 h prior to the addition of LPS. The following groups were then isolated in separate wells: control (manassantin B, 0 ┬ĄM), manassantin B (1, 3 and 10 ┬ĄM), LPS (100 ng/ml) and LPS+ manassantin B (1, 3 and 10 ┬ĄM). The degree of activation of ERK1/2, JNK and p38 MAPK were determined at 30 min and the level of IL-1╬▓ at 24 h after the addition of LPS, respectively.
Cell viability assay
To evaluate the effect of manassantin B on the viability of RAW 264.7 macrophages, the cells were treated with various extract concentrations for 24 h, and then cell viability was evaluated using the methylthiazolyldiphenyl-tetrazolium bromide (MTT) assay. Twenty-four h prior to culture termination, 10 ┬Ąl of the MTT solution (5 mg/ml in PBS, pH 7.4) was added to each well, and the cells were continuously cultured for 4 h. Then 100 ┬Ąl of dimethyl sulfoxide (DMSO) was added for solubilization. Absorbance was measured at 570 nm using an ELISA microplate reader (Molecular Devices, Sunnyvale, CA, USA). Each assay was repeated at least three times.
ELISA
Immunoreactive IL-1╬▓ were quantified using commercially available ELISA kits (R & D Systems, Minneapolis, MN, USA), according to manufacturer's instructions as described previously [17].
Western blot analysis
Western blots to detect levels of phosphorylated and total ERK, JNK and p38 MAPK were performed essentially as previously described [18].
Results
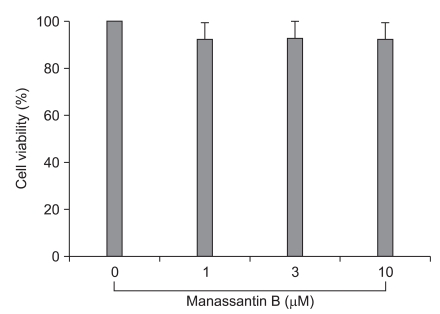
Effect of manassantin B on RAW 264.7 cell viability
The detrimental effects of any compound or plant extract on the cell metabolism need to be determined before the biological activity can be examined. Cell viability was not significantly changed by the presence of manassantin B up to 10 ┬ĄM (Fig. 1).
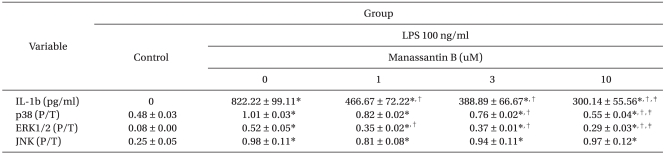
Effects of manassantin B on the expression of pro-inflammatory cytokine IL-1╬▓ in LPS stimulated RAW 264.7 cells
Manassantin B alone did not affect the production of IL-1╬▓ in RAW 264.7 cells over the range of concentrations examined. However, it attenuated an increase in IL-1╬▓ levels induced by LPS in a dose-dependent manner (Table 1).
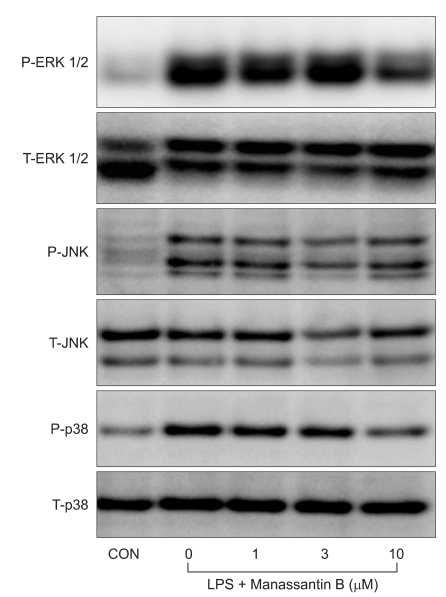
Effects of manassantin B on LPS-induced MAPKs activation
The effects of manassantin B on the phosphorylation of ERK1/2, JNK and p38 MAPK were examined to determine its effect on MAPKs in LPS-stimulated RAW 264.7 cells. In a preliminary study, the maximum MAPKs phosphorylation occurred 30 min after the LPS (100 ng/ml) treatment in RAW 264.7 cells (data not shown). Therefore, the cells were incubated with manassantin B (1, 3, 10 uM) for 1 h and then with LPS (100 ng/ml) for 30 min. Manassantin B attenuated the phosphorylations of ERK1/2 and p38 MAPK in LPS-stimulated RAW 264.7 cells, but had no effect on JNK (Table 1, Fig. 2).
Discussion
In this study, we investigated the effects of manassantin B, an active root extract of Saururus cernuus L, on proinflammatory cytokine production and activation of MAPKs in murine macrophage RAW 264.7 cells stimulated with LPS. Manassantin B attenuated the LPS-induced increase of inflammatory cytokine IL-1╬▓ and phosphorylations of signaling molecules ERK 1/2 and p38 MAPK. These findings suggest that manassantin B has an anti-inflammatory property.
RAW 264.7 murine macrophages are a popular macrophage cell model to examine a variety of inflammatory processes [19-21]. Macrophages are key regulators of inflammation and immunity [22]. Besides being involved in microbial clearance, they play an important role in regulating T cell function, proliferation, regulation of antibody production and production of mediators for cellular immunity. Many of these activities are mediated through pro-inflammatory cytokines [23]. When LPS is presented to Toll-like receptor-4 (TLR-4) in the cell membrane, an interaction via CD 14 and LPS binding protein leads to the activation of MAPKs, which are important for the LPS induced production of a range of inflammatory cytokines, including IL-1, IL-6, TNF-╬▒ and macrophage inflammatory protein (MIP)-2 [24]. These cytokines are essential for the eradication of pathogens. However, excessive release of inflammatory cytokines provokes life-threatening conditions, including sepsis, acute respiratory distress syndrome and multiple organ failure [25].
The MAPK signaling pathways consist of a series of kinases that are activated sequentially and consequently phosphorylate the downstream kinases and transduce extracellular stimuli into intracellular responses. The MAPK family includes ERK, JNK and p38 MAPK. One of the major functions of MAPK is the activation of transcription factors, several of which bind to the promoters of pro-inflammatory cytokines [26]. ERK1/2 regulates TNF-╬▒ and MIP-2 expression and ERK1/2 inhibitor (PD98059) reduces the expression of these cytokines [27,28]. Down-regulation of TLR 4 expression, using RNA interference techniques, decreases MAPKs activation, TNF-╬▒ and MIP-2 expression in RAW 264.7 cells stimulated with LPS [29]. In the present study, manassantin B treatment decreased the phosphorylation of ERK1/2 and p38 MAPK induced by LPS stimulation, but had no affect on JNK phosphorylation, suggesting that its anti-inflammatory activity is related to ERK1/2 and p38 MAPK cascades, but not JNK.
There are some limitations to this study. First, the study is limited in its in vitro character of the data, therefore much remains to be done in vivo studies in terms of safety and optimal dosage. Second, we measured only IL-1╬▓ as a pro-inflammatory cytokine. Potential interactions with other cytokines related with inflammation must be considered. Third, we measured the effects of manassantin B on the phosphorylation of MAPK without inhibitors due to the generalized results of inhibitor study.
In summary, manassantin B inhibits the production of the proinflammatory cytokine IL-1╬▓ and the phosphorylations of ERK 1/2 and p38 MAPK in LPS-stimulated RAW 264.7 murine macrophages. These findings suggest that manassantin B reduces LPS-induced IL-1╬▓ expression through effects on the ERK1/2- and p38 MAPK-mediated pathways in vitro. Thusm manassantin B might be a valuable therapeutic agent in reducing inflammatory response due to systemic inflammatory response syndrome involving sepsis in the clinical setting.